


Estudio in vitro de la influencia de nanopartículas de Au en las líneas celulares HT29 y SPEV
Resumen
Los modelos de cultivo celular son herramientas excelentes para la potencial toxicidad de las nanopartículas y las investigaciones fundamentales en la investigación del cáncer. Por lo tanto, la información sobre la toxicidad potencial de AuNP y los efectos sobre la salud humana es necesaria para el uso de nanomateriales en entornos clínicos. El objetivo de nuestra investigación es examinar los efectos de las AuNP en las líneas celulares de origen epitelial:continuas y oncogénicas. Se utilizaron la línea celular inoculada epitelial de riñón porcino embrionario (SPEV) y la línea celular de carcinoma colorrectal (HT29). En los cultivos de prueba, se evaluaron la proliferación celular, la necrosis / apoptosis y la generación de esferoides multicelulares. Demostramos que las concentraciones de AuNP de 6 a 12 μg / ml redujeron la proliferación de células SPEV y HT29 y aumentaron el número de células en las etapas temprana y tardía de la apoptosis y la necrosis. Se demostró que pequeñas concentraciones de AuNP (1-3 μg / ml) estimulan la formación de esferoides multicelulares por las células HT29 y SPEV. Sin embargo, las concentraciones más altas de AuNP (6-12 μg / ml) tuvieron efectos citotóxicos y anti-cohesivos sobre las células en suspensión. La gran sensibilidad a la acción de las AuNP se demostró mediante la línea de HT29 (6 µg / ml) en comparación con las células SPEV (12 µg / ml). Este estudio experimental del efecto de las AuNP en las líneas celulares SPEV y HT29 justificará su aplicación adicional en el tratamiento contra el cáncer mediado por AuNP.
Antecedentes
La producción y la investigación de las nanopartículas de oro (AuNP) no solo tienen un gran potencial para una amplia aplicación terapéutica del oro [1, 2], sino que también las hacen adecuadas para aplicaciones biomédicas específicas, como las terapias diana [3, 4]. Informes recientes han demostrado que el uso de AuNP ofrece una oportunidad para nuevas terapias antitumorales con un riesgo reducido de desarrollo de resistencias. Por lo tanto, varios estudios han demostrado la actividad antitumoral de las nanopartículas contra el cáncer de mama, hígado, gástrico, colon y pulmón [5, 6].
Se sabe que las nanopartículas (NP) pueden modular el destino celular, inducir o prevenir mutaciones, iniciar la comunicación célula-célula y modular la estructura celular [7, 8]. Además, las AuNP tienen ventajas sobre otras NP metálicas debido a su biocompatibilidad y actividad antitumoral [8,9,10,11,12]. Los efectos citotóxicos y genotóxicos de las AuNP están asociados con su forma, tamaño, carga, concentración, tiempo de interacción, etc. [12, 13, 14]. Por lo tanto, la información sobre su toxicidad potencial y sus efectos en la salud humana es necesaria para el uso de nanomateriales en entornos clínicos.
Actualmente, a pesar del gran éxito de la terapia dirigida, el problema de la administración selectiva de AuNP en el tejido diana sigue sin resolverse. Algunos estudios han observado diferentes tasas de captación de NP por células epiteliales de diferente origen [15, 16]. Sin embargo, faltan investigaciones para explicar este fenómeno a pesar de que pueden ayudar a lograr un objetivo selectivo de tejidos de las AuNP. Las diferencias anatómicas o fisiológicas entre diferentes epitelios podrían explicar las diferencias en las tasas de captación y transporte de AuNP. En particular, la tasa de captación puede verse influenciada por las propiedades de la membrana plasmática de las células y la unión de las nanopartículas a las glicoproteínas y proteoglicanos de la superficie celular, así como por la capacidad de las células para el transporte vesicular [17]. Así, teniendo en cuenta la imposibilidad de interacción exclusivamente selectiva de nanopartículas con células diana, el estudio comparativo de las características de su interacción con células normales y oncogénicas para evitar consecuencias indeseables de la terapia del cáncer es de actualidad [8,9,10].
Aunque los modelos in vivo son valiosos para evaluar la toxicidad biológica de las nanopartículas, los modelos de cultivo celular son muy útiles para estudios fisiológicos y toxicológicos preclínicos. Actualmente, los cultivos celulares se utilizan ampliamente en diversos campos de la biología, la medicina, la medicina veterinaria y la biotecnología. El uso de cultivos celulares permite explorar procesos biológicos que son difíciles y en ocasiones imposibles de estudiar a nivel de organismos. La biotecnología juega un papel importante de los cultivos celulares en la producción de muchas vacunas, sistemas de prueba y sustancias biológicamente activas. Los cultivos celulares se utilizan para diagnosticar enfermedades de diversas etiologías, como objetos de prueba cuando se prueban nuevos agentes farmacológicos, terapéuticos y cosméticos, así como aditivos alimentarios [18].
En este trabajo sobre modelos de cultivo celular, intentamos examinar las características de los efectos de las AuNP de las células epiteliales de origen de línea celular continua y oncogénica. El cultivo en monocapa de células epiteliales SPEV (línea inoculada epitelial de riñón porcino embrionario) y células HT-29 (línea celular de carcinoma de colon) puede considerarse como un modelo de tejidos epiteliales normales y cancerosos cuando se aplica la terapia antitumoral con AuNP. Se emplearon varios ensayos de citotoxicidad tradicionales, que incluyen adhesión, proliferación, necrosis / apoptosis y esferoides multicelulares para validar la citotoxicidad celular de las AuNP.
Métodos
Cultivo de células SPEV
Las células SPEV se cultivaron en matraces de plástico en DMEM (Sigma, EE. UU.) Con FCS al 5% ( v / v ) (HyClone, EE. UU.) Suplementado con penicilina / estreptomicina (PAA, Austria) y anfotericina B (5 μg / ml) (5% CO 2 , 95% de humedad) según lo informado por [19]. La concentración de siembra fue de 0,5 a 2 × 10 4 células / cm 2 . El medio de cultivo se reemplazó cada 2 días. Las células se pasaron al 100% de confluencia [20]. La línea celular SPEV creció y conservó la estructura morfológica inicial de la monocapa durante pases seriados sin evidencia de degeneración celular en cultivo.
Cultivo de células HT29
Las células HT29 se cultivaron en matraces de plástico (Nunc, Dinamarca) en medio RPMI-1640 (Sigma, EE. UU.) Con FCS al 10% ( v / v ) (HyClone, EE. UU.) Suplementado con L-glutamina 2 mM (Sigma, EE. UU.) Y 40 mg / ml de gentamicina (Sigma, EE. UU.) En condiciones estándar (5% CO 2 , 95% de humedad) [21]. La densidad celular óptima fue 0.5–4.0 × 10 4 células / cm 2 . Las células fueron amablemente proporcionadas por el Banco de Líneas Celulares de Tejidos Humanos y Animales del Instituto RE Kavetsky de Patología Experimental, Oncología y Radiobiología NAS de Ucrania.
Manipulaciones con AuNPs
Los AuNP fueron amablemente proporcionados por el Instituto de Bioquímica y Fisiología de Plantas y Microorganismos de la Academia de Ciencias de Rusia. Las AuNP se sintetizaron mediante el método del citrato [22]. El tamaño medio de las nanopartículas fue de 15 nm. La concentración inicial de oro fue de 57 μg / ml. Los resultados de la microscopía electrónica de campo oscuro, la imagen de AuNP de 15 nm y los espectros de extinción de AuNP de 15 nm (b) se muestran en la Fig. 1 (Fig. 1a; nota –– diagrama de la distribución de tamaño) [23]. Las AuNP se introdujeron en las células mediante difusión pasiva a 37 ° C. Las concentraciones investigadas fueron 1, 3, 6 y 12 μg / ml. Las células sin AuNP en las mismas condiciones se tomaron como control.
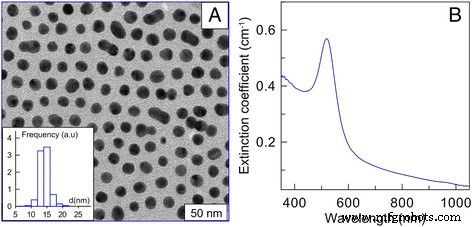
Resultados de la microscopía electrónica de campo oscuro, a imagen de AuNP de 15 nm (nota:diagrama de la distribución de tamaño), b espectros de extinción de AuNP de 15 nm
Células de adhesión y proliferación
El estado morfofuncional de los cultivos celulares se evaluó por las propiedades adhesivas y la actividad proliferativa. Las propiedades adhesivas de las células SPEV y HT29 se evaluaron visualmente usando un microscopio invertido; el número de células adheridas y aplanadas se contó 30, 60, 120, 180 y 1440 min después del inicio del cultivo.
Se estudió la dinámica de proliferación de las células SPEV y HT29 durante 1 a 4 días. Para examinar el incremento del número de células en los cultivos estudiados a los términos de la investigación, se separaron enzimáticamente (1:1 (solución de tripsina al 0,25%:EDTA, PAA)) del plástico y se contó el número de células. El número total de células cultivadas se contó mediante el método tradicional en la cámara de Goryaev.
Procesos apoptóticos / necróticos
Los procesos apoptóticos y necróticos en células SPEV y HT29 expuestas a AuNP se investigaron en 4 días con tintes de Anexina-V (BD, EE. UU.), 7-Amino-Actinomicina (7AAD) (BD) utilizando un FACS Calibur Becton-Dickinson. Los resultados se analizaron con el software WinMDI v.2.8.
Esferoides multicelulares
Los esferoides multicelulares (MS) se generaron para estimar el impacto in vitro de los AuNP sobre el potencial de migración y agregación de las células investigadas. El sistema modelo esferoide (3-D) de células SPEV y HT29 se cultivó mediante el método convencional, que fue informado por [24] y modificado en nuestro laboratorio [25]. Brevemente, la suspensión de células se contó utilizando azul tripán y un número igual de células (5 × 10 4 células / cm 2 ) se sembraron en medio de cultivo completamente suplementado. La generación de EM se logró manipulando el cultivo celular con carboximetilcelulosa (CMC) al 0,24% en placas de 24 pocillos recubiertas con agar al 1% con rotación (80 rpm) durante 24 h. Después de eso, el cultivo de células 3-D se mantuvo en condiciones estándar. Para investigar la dependencia del tamaño y el número de EM de la concentración de AuNP, se generaron EM en presencia de AuNP. Se llevó a cabo un cultivo adicional durante 48 h con rotación constante de placas. En la siguiente etapa, se obtuvieron micro imágenes fotográficas mediante el método de campo oscuro con un microscopio Carl Zeiss Stemi 2000. La morfología de la EM se estudió con la ayuda del programa Axio Vision Release 4.7 (Zeiss). Este programa permite medir las dimensiones geométricas de agregados celulares. Luego, se calculó el volumen de todos los MS que estaban en los archivos. Se usó con la siguiente fórmula: V =0,4 × a × b 2 , donde a y b ––Diámetros geométricos de los MS [24]. Para el análisis estadístico, todos los agregados de células se agruparon por tamaño desde 1 × 10 −4 mm 3 a 1 × 10 −2 mm 3 con incremento de 1 × 10 −3 mm 3 . Se estimó el número de MS y la mediana de los volúmenes de MS para cada grupo.
Análisis estadístico
Un análisis de varianza de un solo factor y la t de Student Se utilizaron pruebas para el procesamiento estadístico de los datos con el paquete de software Statistica 8. El umbral de significancia fue de 0.05. Los resultados se presentan como medias y errores estándar (M ± SE).
Resultados
Efecto de AuNP en la adhesión de células SPEV y HT-29
La adhesión celular es un indicador del estado funcional de las células y es necesaria para un mayor crecimiento del cultivo. Cuando terminó la adhesión, las células se aplanaron y adquirieron la morfología apropiada. Las propiedades adhesivas de las células SPEV se presentan en la Fig. 2.
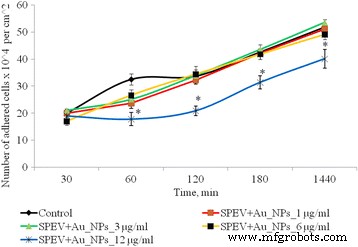
Dinámica de la adhesión de las células SPEV después de la exposición de AuNP, * p ≤ 0.05 es significativo versus con el control
Después de 1 h de cultivo de células SPEV con AuNP a 1, 3 y 6 μg / ml, el número de células adheridas fue menor en comparación con el valor de control. El porcentaje de células aplanadas en muestras con AuNP para estas concentraciones no difirió significativamente del control. La adhesión se ralentizó después de 1 h de incubación con AuNP a 12 µg / ml. El número de células adheridas por centímetro cuadrado se redujo 1,8 veces en comparación con el control. Esta tendencia en la adherencia persistió durante todos los períodos de prueba. Después de 24 h de observación, el número de células adheridas fue 1,3 veces menor que el control. Al mismo tiempo, la incubación de AuNP a pequeñas concentraciones (1 y 3 μg / ml con células tumorales (HT29) no tuvo un efecto significativo sobre la cantidad de células adhesivas. El aumento de la concentración de AuNP a 6 y 12 μg / ml conduce a una disminución de la número de células tumorales en la fracción adhesiva en 1,16 y 1,28 veces, respectivamente, (Fig. 3). Los datos obtenidos pueden verse influenciados por varios procesos. Uno es el efecto citostático / citotóxico de las AuNP sobre la fracción de adhesión tanto del tumor como líneas celulares embrionarias, que conduce a la muerte celular, transición a la apoptosis o necrosis. El otro proceso es la reducción de la adhesión celular, bajo la influencia de AuNPs y la transferencia de células a la fracción de suspensión. Cabe destacar que ambos procesos se pueden realizar simultáneamente, y cada uno puede contribuir a la disminución del número de células vivas en la fracción de adhesión.
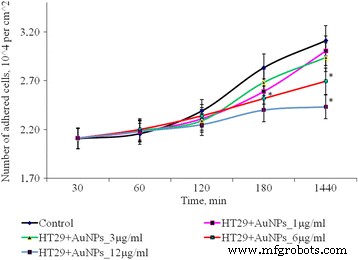
Dinámica de la adhesión de las células HT29 después de la exposición de AuNP, * p ≤ 0.05 es significativo versus con el control
Efecto de las AuNP en la proliferación de células SPEV y HT-29
Se estudió el efecto de las AuNP en el rango de concentración de 1 a 12 μg / ml sobre los procesos proliferativos en cultivos de células SPEV (Fig. 4). En los días 2-4 de cultivo con AuNP a 1, 3 y 6 μg / ml, el número de células no difirió significativamente del control. El día 4 de cultivo con AuNP a 3 y 6 μg / ml, este índice disminuyó 1,15 y 1,23 veces, respectivamente, en comparación con el control. Se observó una reducción en el número de células en 1,5 veces (días 2 y 3) y en 1,15 veces el día 4 de cultivo con AuNP a 12 µg / ml en cultivo SPEV frente al control. Por lo tanto, la concentración de AuNP, 12 μg / ml, ralentizó el crecimiento celular dentro del período de tiempo observado.
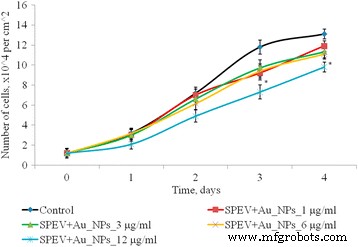
Proliferación de células SPEV después de la exposición de AuNP, * p ≤ 0.05 es significativo versus con el control
El efecto de AuNP a concentraciones de 1 a 12 μg / ml sobre el número de células HT 29 en un cultivo monocapa se muestra en la Fig. 5. Durante los primeros 3 días de incubación, el número de células en el control y en presencia de AuNPs no fue estadísticamente diferente. En el cuarto día de cultivo, se observó una disminución dependiente de la dosis del número de células en cultivo 2D. Entonces, después de 4 días de cultivo, para concentraciones bajas de AuNP (1 y 3 μg / ml), el número de células HT 29 no es significativamente diferente en comparación con el control. Pero a concentraciones más altas de AuNP (6-12 μg / ml), el número de células HT29 fue menor que el control en 1,33 y 1,44 veces, respectivamente.
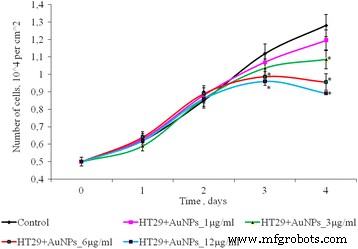
Proliferación de células HT29 después de la exposición de AuNP, * p ≤ 0.05 es significativo versus con el control
Efecto de AuNP en procesos apoptóticos / necróticos en células SPEV y HT-29
Se cultivaron células SPEV y HT-29 en presencia de AuNP durante 4 días en condiciones estándar. El cultivo de células SPEV y HT29 con AuNP a 1 y 3 μg / ml y los índices de procesos apoptóticos / necróticos no difirieron significativamente del control (Tablas 1 y 2).
El cultivo con AuNP a 6–12 μg / ml aumentó el porcentaje de células anexina V + / 7AAD +, anexina V- / 7AAD + y anexina V + / 7AAD y redujo el porcentaje de células vivas. El número de anexina V + / 7AAD + Las células SPEV fueron más altas que el valor de control en 7.8 ± 0.7% ( p ≤ 0,05) con 12 μg / ml de AuNP. El número de anexina V + / 7AAD + Las células HT 29 fueron más altas que el valor de control en un 3,2 ± 0,4% ( p ≤ 0,05) con 6 μg / ml de AuNP y en un 4,8 ± 0,6% ( p ≤ 0,05) con 12 μg / ml de AuNP.
Efecto de AuNP en la generación de esferoides multicelulares a partir de células SPEV y HT29
Para determinar la dependencia del tamaño y el número de esferoides multicelulares (MS) de la concentración de AuNP, se generaron MS a diversas concentraciones de AuNP durante 48 h. Nuestros datos demostraron la capacidad de variedad de las células HT29 y SPEV para formar esferoides multicelulares en las mismas condiciones del microambiente (Figuras 6 y 7).
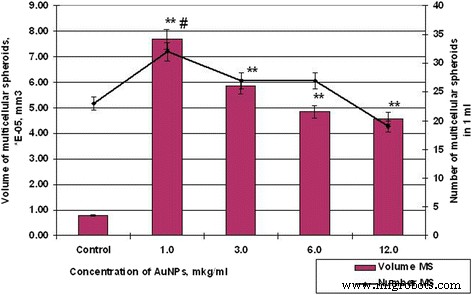
Número y volumen de SPEV de células MS después de la incubación con AuNP, # p ≤ 0,01 (para número de EM); ** p ≤ 0.01 (para volumen de MS)
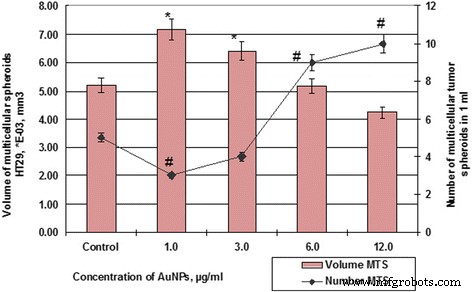
Número y volumen de células de MS HT29 después de la incubación con AuNP. # p ≤ 0,01 (para número de EM); ** p ≤ 0.01 (para volumen de MS)
Entonces, si las muestras de control de células HT29 durante 48 h formaron esferoides en un volumen promedio de 5.19 × 10 −3 mm 3 , el esferoide de volumen promedio de las células SPEV fue de 0,79 × 10 −5 mm 3 . Al mismo tiempo, la influencia de AuNP en las células HT29 y SPEV tuvo la misma tendencia. La presencia de AuNP en el microambiente celular estimuló la formación de esferoides multicelulares en ambos cultivos. Por lo tanto, cuando la concentración de AuNP fue de 1 y 3 μg / ml, el volumen de MS para SPEV aumentó en 9,7 y 7,4 veces, respectivamente, en comparación con el control (Fig.6), las mismas concentraciones de AuNP también estimularon un volumen creciente de MS para HT29 en 1,4 y 1,2 veces, respectivamente (Fig. 7).
El aumento adicional de la concentración de AuNP condujo a una disminución del volumen medio de EM en ambos cultivos. La elevación en la concentración de AuNP de 1 a 12 μg / ml disminuyó el volumen de HT29 MS de 7,18 × 10 −3 mm 3 hasta 4,24 × 10 −3 , en 1,69 veces, según el control. En cuanto a SPEV, cuando la concentración de AuNP aumentó de 1 a 12 μg / ml, el volumen de MS disminuyó de 7,69 a 4,58 × 10 −5 mm 3 , en 1,68 veces, según control. Sin embargo, el aumento de la concentración de AuNP coincide con la reducción del volumen de EM y se correlaciona con el aumento del número de esferoides en el cultivo de células HT29 (Figuras 6 y 7). El número de HT29 MS aumentó de 3 a 10 por campo de visión a una concentración de AuNP de 1 a 12 μg / ml. Al mismo tiempo, el número de SPEV MS disminuyó de 32 a 19, respectivamente.
Los datos obtenidos (Figuras 6 y 7) demuestran que las AuNP son capaces de influir en las interacciones cohesivas en el sistema de célula a célula. Nuestros datos muestran que pequeñas concentraciones de AuNP (1-3 μg / ml) estimularon la formación de esferoides multicelulares de células embrionarias y tumorales. Sin embargo, las concentraciones más altas de AuNP (6-12 μg / ml) tuvieron efectos citotóxicos y anti-cohesivos sobre las células en suspensión. Este proceso contribuyó a la formación de un mayor número de HT29 MS con la disminución del volumen medio. En cuanto a SPEV, la alta concentración de AuNP puede tener un efecto citostático que reduce el número de células en la fracción adhesiva y el número de MS en suspensión. Anteriormente, los autores informaron que las nanopartículas de carbono reducen la adhesión de las células al sustrato, estimulan la transferencia celular a la suspensión y conducen a la formación de esferoides multicelulares [25, 26]. En la literatura, hay datos sobre la capacidad de AuNP para romper la estructura de los microfilamentos de actina / miosina y disminuir la proliferación, adhesión y diferenciación celular [27]. Nuestros datos confirmaron esta suposición.
Discusión
Se evaluaron los efectos de las AuNP sobre la proliferación, necrosis / apoptosis y formación de esferoides multicelulares de las células epiteliales continuas y de origen de la línea celular oncogénica. Se demostró que las AuNP a 6-12 μg / ml redujeron el número de células SPEV y HT29 y aumentaron el número de células en las etapas temprana y tardía de la apoptosis y la necrosis. Las pequeñas concentraciones de AuNP estimulan la formación de esferoides multicelulares por las células HT29 y SPEV. Sin embargo, concentraciones más altas de AuNP tuvieron efectos citotóxicos y anti-cohesivos sobre las células en suspensión. La gran sensibilidad a la acción de las AuNP se demostró mediante la línea de HT29 (6 μg / ml) en comparación con las células SPEV (12 μg / ml).
Los efectos de las AuNP sobre la morfología celular y el citoesqueleto solo han recibido más atención recientemente, y el mecanismo subyacente y las consecuencias futuras no se han investigado en profundidad [28, 29, 30]. En este sentido, es importante que todos los nuevos tipos de AuNP evalúen su vía de captación endocítica y su localización intracelular en función del tiempo. Para diferentes tipos de AuNP, se ha descrito que los efectos dependen de la concentración de AuNP intracelular y son transitorios, donde después de divisiones celulares recurrentes, las concentraciones de AuNP intracelulares disminuyen exponencialmente y los efectos ya no se observan. Además, se debe evaluar el posible escape endosómico de las AuNP. Como se ha descrito que los defectos del citoesqueleto dependen claramente de las concentraciones de AuNP, se debe probar una amplia gama de concentraciones de partículas para intentar evaluar la capacidad de carga celular máxima sin ningún efecto. Además, como el citoesqueleto también participa en muchas vías de señalización intracelular, queda por investigar si la alteración citoesquelética inducida por AuNP conduce a efectos secundarios [31].
Como las NP tienen ciertas dimensiones físicas, el volumen intracelular que ocupan puede provocar alteraciones en la morfología celular o afectar la estructura de la red del citoesqueleto celular [28, 29, 31]. Los efectos posteriores también pueden deberse a las altas exigencias de la postura NP sobre la forma endocítica celular. Se ha descrito que las AuNP tienen un efecto profundo en la morfología de varios tipos de células, como las células pulmonares de carcinoma humano A549 [32]. También se ha descrito que las AuNP tienen un efecto dependiente de la concentración sobre las fibrillas de actina de los fibroblastos dérmicos humanos [33, 34]. Mironava y col. [35, 36] mostró además que los filamentos del citoesqueleto se interrumpían en función del tiempo de exposición de AuNP, la concentración y el tamaño de las NP, aunque los niveles de expresión de la proteína actina o β-tubulina no se vieron afectados.
El tipo de célula utilizado también es de gran importancia, ya que diferentes tipos de células, incluso cuando están estrechamente relacionados, pueden reaccionar de manera muy diferente para el mismo tipo de nanomateriales [37, 38]. Preferiblemente, los tipos de células que están más involucrados en las (futuras) aplicaciones biomédicas de las NP deberían probarse (por ejemplo, células epiteliales, endoteliales), o células múltiples que se derivan de las diferentes capas germinales. Al investigar los efectos citotóxicos, debe minimizarse el uso de tipos de células cancerosas, ya que pueden dar lugar a resultados aberrantes [39]. Las células cancerosas tienen varias características específicas y vías de señalización intracelular alteradas que están destinadas a regular positivamente la proliferación y mantener la viabilidad celular, lo que las hará menos propensas a algunos efectos citotóxicos mediados por NP.
En nuestra opinión, la unión de las AuNP a los grupos funcionales de la superficie (p. Ej., Proteínas transmembrana) de las células puede ser reversible o irreversible, lo que da lugar a lesiones estructurales temporales o permanentes [40, 41]. Se pueden percibir las posibles implicaciones de los cambios en las propiedades biomecánicas (p. Ej., Dureza y elasticidad), adhesividad y propiedades eléctricas superficiales de las células. Por tanto, es probable que los cambios en la dureza o la elasticidad influyan en la flexibilidad estructural de la superficie, la producción de energía mecánica para la división celular y la motilidad celular. En cuanto a la adhesividad, el microambiente celular normalmente está compuesto por una matriz extracelular con moléculas específicas que permiten que las células se adhieran a su entorno [42]. Sin duda, la carga superficial juega un papel importante en las interacciones entre las células y su entorno.
Los otros autores también informaron de que las NP se localizan preferentemente en las mitocondrias y causan estrés oxidativo, además de potenciar el daño estructural [40]. Un artículo reciente de Pan et al. describe que las AuNP de 1,4 nm inducen necrosis a través del estrés oxidativo y el daño mitocondrial en las células Hela [43]. La acumulación de nanopartículas en el medio celular tras la biodegradación no es segura porque puede alterar los orgánulos e incluso causar mutaciones genéticas.
Los cambios que ocurren en las células durante la apoptosis son similares para la mayoría de los tipos de células. En las células apoptóticas, hay cambios en la composición de lípidos de la membrana plasmática:la fosfatidilserina se transfiere desde la parte citoplásmica de la bicapa al lado externo, lo que provoca la activación de la cascada de caspasas, la condensación de cromatina y el trastorno de la cadena de transporte de electrones en las mitocondrias y, finalmente, deteniendo la síntesis de ATP. La muerte celular programada puede desencadenarse por estímulos fisiológicos mediados por receptores resultantes de trastornos genéticos, exposición a factores químicos o físicos, así como por otros cambios en las células. Observamos que este efecto es con 6-12 μg / ml de AuNP.
Los agregados multicelulares (esferoides, cuerpo embrioide) representan un nivel intermitente entre las células en crecimiento monocapa y el cultivo de tejidos. Los esferoides son un modelo objetivo del crecimiento y organización celular tridimensional, las interacciones célula a célula y la influencia de las condiciones microambientales, por ejemplo, la concentración de AuNP, sobre la intensidad de la proliferación, así como sobre la adhesividad celular y la formación de microagregados. La formación de EM es un método de cultivo bien establecido tanto para el tumor como para las líneas celulares embrionarias [24, 44, 45]. En nuestro trabajo, la formación y el crecimiento de esferoides se logra mediante la adición de CMC como parte de la matriz extracelular artificial y el recubrimiento superficial con agar al 1% que inhibe la adhesión celular a la superficie y estimula la agregación celular. En estas condiciones, el cultivo de EM puede realizarse hasta que los agregados formen necrosis central, debido al crecimiento de masa celular limitado o la diferenciación espontánea de las células embrionarias.
En la literatura, hay información sobre la interacción AuNP con la línea celular de cáncer de colon y líneas celulares embrionarias [46, 47]. Según estos datos, la exposición incluso a concentraciones muy bajas de AuNP puede tener un efecto dañino sobre las células precursoras neuronales embrionarias humanas y HT29 al hacer hincapié en la proliferación celular, la diferenciación y la muerte celular apoptótica.
Hay datos publicados de que el efecto de las AuNP se basa en la acumulación de la fase G0 / G1, la depleción de la fase S y G2 / M, así como en la reducción de los niveles de ATP en las células del carcinoma escamoso oral humano (HSC-3) [48]. La regulación del ciclo celular puede abordarse mediante la violación de los contactos focales de las células con el sustrato y la transferencia celular a la fracción suspendida en cultivo 2D y la inhibición de los contactos célula a célula en la unión gap en el cultivo 3D [48,49,50]. Debido al tamaño nanométrico de las AuNP (cerca de 15 nm), no pueden ser centros de cohesión para las células. Al mismo tiempo, la intercalación de AuNP en la membrana celular [51], la influencia en el potencial zeta de AuNP de la membrana celular [32] y la influencia en la formación de contactos de célula a célula / célula a superficie obviamente pueden desencadenar el mecanismo de necrosis / apoptosis , efecto citotóxico y detención del ciclo celular. La violación de los contactos focales de las células con el sustrato y la transferencia celular a la fracción suspendida es una forma de regulación del ciclo celular [48, 49]. Pequeñas concentraciones de AuNP no ejercieron ningún efecto citotóxico estadísticamente significativo en las células. Sin embargo, concentraciones más altas de AuNP tuvieron efectos citotóxicos y anti-cohesivos sobre las células en suspensión. Este proceso contribuyó a la formación de un mayor número de MS con la disminución del volumen promedio. Suponemos que AuNP se encaja en contactos cohesivos de células y las compromete. Por lo tanto, nuestros experimentos sobre los efectos de las AuNP en las líneas celulares SPEV y HT29 respaldan su aplicación adicional en el desarrollo de terapias contra el cáncer mediadas por AuNP.
Aunque serán necesarios estudios futuros para confirmar los efectos anticancerígenos en los estudios con animales in vivo. Sin embargo, nuestra profunda convicción es que si conocemos la naturaleza de la sustancia y su posible influencia negativa, podemos evitar los efectos perjudiciales de las AuNP y aprovechar su potencial biotecnológico positivo. Nuestra investigación podría aplicarse de manera bastante confiable en materiales efectivos para el contexto del tratamiento contra el cáncer con la máxima ventaja para la medicina.
Conclusiones
Nuestros resultados apoyan la idea de que las AuNP inducen citotoxicidad dependiente de la dosis en células SPEV y HT29. Además, este informe demuestra por primera vez que las AuNP de 15 nm en concentraciones de 6 a 12 μg / ml redujeron la proliferación de células SPEV y HT29 y aumentaron el número de células en las etapas temprana y tardía de la apoptosis y la necrosis. Además, se demostró que pequeñas concentraciones de AuNP (1-3 μg / ml) estimulan la formación de esferoides multicelulares. Sin embargo, concentraciones más altas de AuNP tuvieron efectos citotóxicos y anti-cohesivos sobre las células en suspensión. La gran sensibilidad a la acción de las AuNP se demostró mediante la línea de HT29 (6 μg / ml) en comparación con las células SPEV (12 μg / ml).
Nanomateriales
- Nanopartículas de oro para sensores de quimioterapia
- Nanopartículas de oro multifuncionales para aplicaciones terapéuticas y diagnósticas mejoradas:una revisión
- Nanopartículas para la terapia del cáncer:avances y desafíos actuales
- Preparación y propiedades magnéticas de nanopartículas de espinela de FeMn2O4 dopadas con cobalto
- Preparación de nanopartículas de mPEG-ICA cargadas con ICA y su aplicación en el tratamiento del daño celular H9c2 inducido por LPS
- Bioseguridad y capacidad antibacteriana del grafeno y el óxido de grafeno in vitro e in vivo
- Síntesis y rendimiento in vitro de nanopartículas de hierro-platino recubiertas de polipirrol para terapia fototérmica e imágenes fotoacústicas
- Promoción del crecimiento celular SH-SY5Y mediante nanopartículas de oro modificadas con 6-mercaptopurina y un péptido penetrante de neuronas
- Influencia de la rigidez elástica y la adherencia de la superficie en el rebote de nanopartículas
- Una metodología de dos pasos para estudiar la influencia de la agregación / aglomeración de nanopartículas en el módulo de Young de nanocompuestos poliméricos
- Comparación entre el ácido fólico y la funcionalización basada en péptidos gH625 de nanopartículas magnéticas de Fe3O4 para una internalización celular mejorada