


Las nanopartículas modificadas con hGC33 y cargadas con sorafenib tienen un efecto antihepatoma sinérgico al inhibir la vía de señalización Wnt
Resumen
La administración de inhibidores específicos de tumores es un desafío en el tratamiento del cáncer. Las nanopartículas modificadas con anticuerpos pueden administrar sus fármacos cargados a las células tumorales que sobreexpresan antígenos específicos asociados a tumores. Aquí, construimos nanopartículas de polímero de polietilenglicol-b-PLGA cargadas con sorafenib modificadas con anticuerpo hGC33 contra glypican-3 (GPC3 +), una proteína de membrana sobreexpresada en el carcinoma hepatocelular. Descubrimos que los NP modificados con hGC33 (hGC33-SFB-NP) tenían como objetivo GPC3 + Las células del carcinoma hepatocelular (HCC) al unirse específicamente a GPC3 en la superficie de las células de HCC, inhibieron la transducción de señales inducida por Wnt e inhibieron las células de HCC en G0 / 1 al regular a la baja la expresión de ciclina D1, atenuando así la migración de las células de HCC al inhibir las células epiteliales. transición mesenquimal. hGC33-SFB-NP inhibió la migración, la progresión del ciclo y la proliferación de células de HCC al inhibir la vía Ras / Raf / MAPK y la vía Wnt en conjunto con moléculas GPC3, respectivamente. hGC33-SFB-NP inhibió el crecimiento del cáncer de hígado in vivo y mejoró la tasa de supervivencia de los ratones portadores de tumores. Llegamos a la conclusión de que hGC33 aumenta la orientación de SFB-NP a las células HCC. hGC33-SFB-NP inhibe sinérgicamente la progresión del HCC al bloquear la vía Wnt y la vía Ras / Raf / MAPK.
Introducción
Glypican3 (GPC3) es un proteoglicano de heparán sulfato que se expresa en las superficies celulares a través de un mecanismo que implica anclas de glicerofosfatidilinositol [1]. Aunque GPC3 se expresa en una variedad de tejidos durante el desarrollo, su expresión se inhibe al menos parcialmente en la mayoría de los tejidos adultos por la metilación del ADN en la región promotora [2]. Sin embargo, la proteína GPC3 se sobreexpresa en aproximadamente el 70% de los pacientes con carcinoma hepatocelular (HCC) [3, 4] y estimula la transducción de la señal Wnt clásica [5] a través de la interacción con el ligando Wnt, que promueve la unión de Wnt / frizzled para promover el crecimiento del HCC [6] . La activación de la vía de señalización clásica de Wnt es uno de los eventos más frecuentes asociados con la transformación maligna del CHC [7, 8]. Basándonos en la capacidad de GPC3 para aumentar la señalización de Wnt, planteamos la hipótesis de que la sobreexpresión de GPC3 promueve el crecimiento de HCC al estimular la vía clásica de Wnt.
Anticuerpo monoclonal terapéutico que reconoce un epítopo en la porción C-terminal de GPC3 (524-563) (hGC33), que reconoce el epítopo C-terminal de GPC3 (524-563), inhibe el crecimiento tumoral en xenoinjertos subcutáneos de HepG2 y Huh- 7 en ratones [9, 10]. La hGC33 se ha humanizado mediante el trasplante complementario de la región de decisión, y su efecto anticanceroso es tan eficaz como la hGC33 para el xenotrasplante de HepG2, y la hGC33 se ha utilizado en ensayos clínicos [11]. Estos resultados sugieren que hGC33 tiene una importante actividad antitumoral, y el tratamiento anti-GPC3 inhibirá directamente la proliferación y / o supervivencia de las células de HCC al bloquear Wnt y / u otras vías de señalización.
Sin embargo, debido a la compleja patogénesis del CHC, la eficacia del bloqueo de GPC3 solo es limitada [12]. Por lo tanto, explorar el mecanismo detallado de la patogénesis del CHC e identificar biomarcadores prometedores para el diagnóstico y pronóstico del CHC puede ayudar a proporcionar dianas terapéuticas eficaces y mejorar el pronóstico de los pacientes. Se ha propuesto que los anticuerpos anti-GPC3 aumentan la sensibilidad del CHC a los agentes quimioterapéuticos [13]. La actividad antitumoral de hGC33 en combinación con fármacos quimioterapéuticos estándar se ha evaluado recientemente [14]. El sorafenib es un nuevo fármaco oral dirigido contra el HCC que puede inhibir directamente la proliferación de células tumorales al bloquear la vía de señalización celular mediada por RAF / MEK / ERK para frenar el crecimiento tumoral [15, 16]. Sin embargo, la distribución no dirigida de sorafenib in vivo y las señales Wnt activadas de forma anormal en el HCC limitan la eficacia del fármaco y aumentan sus efectos secundarios [14, 15, 16]. Wnt se une a los receptores de la familia Frizzled para activar la transducción de señales intracelulares que regula la proliferación celular, la apoptosis y la migración celular, y causa resistencia a los fármacos en muchos tumores, como el HCC [17,18,19].
En los modelos de xenotrasplante de HepG2, la combinación de hGC33 y sorafenib es más eficaz para inhibir el crecimiento tumoral que el sorafenib solo [15], y la administración de fármacos con nanoportadores de polímero ha recibido mucha atención en el tratamiento del cáncer. Los nanoportadores cargados con fármacos anticancerígenos pueden prevenir la distribución inespecífica y la degradación inespecífica in vivo, mejorar la biodisponibilidad del fármaco y la orientación antitumoral y simplificar la evaluación de la farmacocinética y el tratamiento [15, 16]. En una variedad de nanopartículas a base de polímeros, las formulaciones a base de poli (ácido láctico coglicólico) (PLGA) se consideran portadores de fármacos ideales y seguros [17, 18]. En este sentido, el poli (etilenglicol) - b -poli (d, l-lactida-co-glicólido) (PEG- b -PLGA) se basan en polietilenglicol y copolímeros de PLGA, que son seguros y no tóxicos después de la hidrólisis, y han sido aprobados por la Administración de Drogas y Alimentos de los Estados Unidos [19,20,21,22]. Por lo tanto, inyección intravenosa con nanoportador de PEG- b -El copolímero PLGA es una estrategia prometedora para lograr una entrega dirigida y mejorar la eficacia. Además, la aplicación del anticuerpo específico hGC33 contra moléculas GPC3 en la membrana celular de HCC no solo puede mejorar la administración de nanofármacos dirigidos in vitro e in vivo [18, 19], sino que también bloquea Wnt y / u otras vías de señal conectadas con GPC3, inhiben la proliferación y / o supervivencia de las células cancerosas, y puede lograr una actividad antitumoral sinérgica.
En este estudio, exploramos si el copolímero PEG- b modificado con anticuerpo hGC33 -Nanopartículas de PLGA pueden facilitar la administración de sorafenib (hGC33-SFB-NP) a HCC in vivo e in vitro, y mejorar la eficiencia del tratamiento de HCC a través de la selección activa de HCC para cambiar la farmacocinética del fármaco. De acuerdo con el tamaño de partícula, el potencial zeta, la morfología de las partículas, la eficacia de atrapamiento del fármaco, la capacidad de carga del fármaco y la liberación del fármaco in vitro, se caracterizó exhaustivamente el NP objetivo. La capacidad de direccionamiento in vitro se caracteriza por la captación celular de las células del hepatoma HepG2. La biodistribución y el efecto terapéutico sinérgico de hGC33-SFB-NP sobre el HCC se evaluaron comparando sorafenib y SFB-NP. Nuestros resultados demostraron que hGC33-SFB-NP puede apuntar a GPC3 + HCC. Puede inhibir la progresión del ciclo celular, la proliferación celular y la invasión tumoral al inhibir las vías de señal Wnt y Ras / Raf / MAPK e inhibir sinérgicamente la progresión del cáncer de hígado.
Materiales y métodos
Materiales
El anticuerpo hGC33, bromuro de 3- (4,5-dimetiltiazol-2-il) -2,5-difeniltetrazolio (MTT), 4 ', 6-diamidino-2-fenilindol, diclorhidrato (DAPI), 5,5', 6 , Yoduro de 6'-tetracloro-1,1 ', 3,3'-tetraetil-bencimidazolil-carbocianina (JC-1) y dimetilsulfóxido (DMSO) se obtuvieron de Sigma-Aldrich, Inc. (St. Louis, MO). Clorhidrato de 1- (3-dimetilaminopropil) -3-etilcarbodiimida y N -hidroxisuccinimida se obtuvieron de Qiyun Biotech (Guangzhou, China). El kit de cuantificación de proteínas de ácido bicinconínico (BCA), cumarina-6, y el kit de detección de apoptosis Annexin V-FITC / PI se adquirieron de Beyotime Biotechnology (Shanghai, China). PEG- b -Copolímero dibloque PLGA-maleimida (mal-PEG- b -PLGA; 25 000-30 000 Da, PLGA, LA:GA, p / p; PEG, 13-15%) se adquirió de Polyscitech (West Lafayette, IN, EE. UU.). Kit de anticuerpos PI3K (9655 #), kit de anticuerpos p-Akt (9916 #), kit de anticuerpos mTOR (9964 #), kit de anticuerpos de la familia Bcl-2 (9942 #), kit de anticuerpos de apoptosis (9915 #) y anti- los anticuerpos de conejo y anti-ratón se adquirieron de Cell Signal Technology (Danvers, MA, EE. UU.); La ciclina B1 y la quinasa dependiente de ciclina se adquirieron de Abcam Biological Technology (EE. UU.). Los anticuerpos contra fosfo-Rb, ciclina D1, punto de control quinasa 1 (CHK1), P53, gen 1 de susceptibilidad al cáncer de mama fosforilado (p-BRCA1), RAD51, citocromo C y metaloproteinasa de matriz (MMP2 y MMP9) se compraron a Abcam (EE. UU.) . Todos los demás productos químicos, reactivos y disolventes de grado analítico se obtienen de proveedores estándar y se utilizaron sin purificación adicional.
Células y animales
La línea celular de HCC HepG2 (obtenida de la colección de cultivos tipo estadounidense (Manassas, VA, EE. UU.) Se cultivó en medio DMEM (Invitrogen, Carlsbad, CA, EE. UU.) Suplementado con FBS al 10% (HyClone, Logan, UT, EE. UU.) Y 80 U / ml de penicilina y 80 μg / ml de estreptomicina en atmósfera humidificada de 5% de CO 2 a 37 ° C. Nanjing Junke Biotechnology Co., Ltd. (China) proporcionó ratones desnudos BALB / C, con un peso de 20 a 22 g (5 a 6 semanas). Se criaron ratones desnudos BALB / C en una habitación SPF. Todo el cuidado y tratamiento de los animales se llevó a cabo de acuerdo con los requisitos de cuidado animal de la Universidad de Ciencia y Tecnología de Anhui. Todos los protocolos experimentales han sido revisados y aprobados por el comité de ética de experimentos con animales de la Universidad de Ciencia y Tecnología de Anhui (número de aprobación:2019dw013).
Preparación de NP
Para preparar NP, mal-PEG- b Se pesaron -PLGA y SFB o cumarina-6 y se disolvieron en la fase orgánica (diclorometano / acetona 3:2 v / v). La solución se añadió a una solución de alcohol polivinílico (PVA) (5% p / v) gota a gota con agitación continua. La mezcla se sometió a ultrasonidos de forma intermitente con un sonicador de sonda (potencia de salida de 550 W, 8 veces) en hielo para crear una emulsión de aceite y agua. La emulsión se añadió a una solución de PVA (1% p / v) con agitación magnética. Las NP de SFB y cumarina-6 se recogieron mediante centrifugación a 8000 rpm durante 30 min y se lavaron tres veces en agua Milli-Q.
Para generar hGC33-NP mediante enlaces tioéter formados por la reacción de maleimida con residuos de sulfhidrilo libres en el anticuerpo hGC33, el anticuerpo hGC33 se mezcló con NP funcionalizadas con maleimida en una proporción molar de 5:1 (hGC33:mal-PEG- b -PLGA) y se incubó a 4 ° C durante 16 h con agitación continua. La hGC33 se conjugó con las NP mediante la reacción de los grupos sulfhidrilo del anticuerpo hGC33 con los grupos maleimida de las cadenas de PEG. Los anticuerpos no conjugados se eliminaron pasándolos a través de columnas de Sepharose CL-4B. La conjugación eficaz de proteínas se confirmó con un kit BCA (Thermo Fisher Scientific, Waltham, MA, EE. UU.).
Caracterización de nanopartículas (NP)
Morfología, tamaño de partícula, eficiencia de encapsulación (EE) y estabilidad de NP
La morfología de las NP se evaluó con microscopía electrónica de transmisión (TEM, H-600; Hitachi, Tokio, Japón). Las hGC33-NP sin fármaco y las NP sin fármaco se registraron mediante espectrofotómetro FTIR (Thermo Nicolet, Madison, WI, EE. UU.) Utilizando bromuro de potasio. El tamaño medio de partícula y el potencial zeta de las NP se caracterizaron con un Malvern Zetasizer ZEN3600 Nano ZS (Malvern Instruments, Malvern, Reino Unido) a 20 ° C. La eficacia de encapsulación del fármaco (EE) y la eficacia del contenido de carga del fármaco (LC) se evaluaron mediante ultrafiltración. Las muestras se cargaron en un dispositivo de ultrafiltración (100 kMWCO; Sartorius, Göettingen, Alemania) y se centrifugaron a 8000 rpm durante 25 min a 4 ° C para eliminar el fármaco libre. Se disolvió el mismo volumen de cada muestra en acetonitrilo para confirmar la cantidad total de fármaco. La concentración se midió mediante cromatografía líquida de alta resolución. La longitud de onda de absorción fue de 266 nm. Se utilizó la siguiente fórmula para calcular el EE del fármaco (%) de los NP:(peso del fármaco atrapado / peso total del fármaco) × 100%. La LC (%) se calculó como (peso de fármaco encapsulado / peso de NP) x 100%. Para comprender la estabilidad de las NP a temperatura ambiente, se evaluó el cambio en el tamaño de las NP mediante dispersión dinámica de luz (DLS) en puntos de tiempo predeterminados (0,5, 1, 2, 4, 8, 12 h, 16 h, 20 h y 24 h). h) a 25 ° C.
Ensayo de absorción celular y liberación de fármaco in vitro
El fármaco de las NP se investigó utilizando bolsas de diálisis con un límite de peso molecular de 10 kDa. Brevemente, se cargó 1 ml de NP en una bolsa de diálisis (MWCO 8.000–10.000 Da; Spectrum Labs Inc., CA, EE. UU.). Las bolsas de diálisis se sumergieron en PBS y se agitaron con un agitador magnético a 25 ° C. Los perfiles de liberación de fármacos de las NP se midieron en 100 ml de solución salina tamponada con fosfato 0,2 M (PBS; pH =7,4) durante 7 días. La concentración de fármaco en las muestras se midió con cromatografía líquida de alta resolución. En estudios posteriores, se usó hGC33-cumarina 6-NP que tenía el mismo tamaño de partícula que hGC33-SFB-NP para evaluar el direccionamiento de hGC33-SFB-NP. HepG2 (GPC3 + ) y Li-7 (GPC3 - ) se incubaron con hGC33-cumarina 6-NP durante 0,5, 2 o 4 ha 37 ° C en CO 2 al 5% respectivamente. Las células cocultivadas se lavaron y fijaron con formaldehído al 4% durante 10 min; los núcleos celulares se tiñeron con 5 μg / mL Hoechst 33,342 durante 15 min para localizar NP intracelulares. La microscopía confocal para analizar imágenes de nanopartículas intracelulares se analizó con microscopía confocal (Olympus FV1000; Olympus Corporation, Tokio, Japón).
Efecto celular in vitro
La citotoxicidad de hGC33 libre (Ab), SFB libre, hGC33-null-NP o hGC33-SFB-NP se determinó usando un ensayo MTT. HepG2 (GPC3 + ) celdas y Li-7 (GPC3 - ) se sembraron células en fase logarítmica en una placa de 96 pocillos a una densidad de 4000 células por pocillo seguido de incubación durante 48 ha 37 ° C en 5% de CO 2 . Las células se trataron con hGC33, SFB libre, hGC33-null-NP o hGC33-SFB-NP durante 48 ha 37 ° C en 5% CO 2 . Después del co-cultivo durante un tiempo definido, se determinó la actividad de proliferación celular mediante el ensayo MTT como se describe [20]. La absorbancia de cada pocillo se midió a 490 nm y se calculó la mitad del valor de concentración inhibitoria máxima (IC 50) con SPSS 17.0.
Medición de la capacidad de invasión celular
Las células en fase de crecimiento logarítmico se sembraron en placas de 6 pocillos a una densidad de 5 × 10 4 células / pocillo, raspado con un cabezal de succión y reemplazado con medio de cultivo sin suero. La cicatrización de la herida se registró a las 0 h, 24 hy 48 h en el grupo de control y en el grupo experimental. Al mismo tiempo, se inoculó el mismo número de células en cámaras Transwell y se trató con hGC33 libre, SFB libre, hGC33-null-NP o hGC33-SFB-NP. Se añadieron quinientos microlitros de medio FBS al 10% en la cámara inferior. Después de 24 h de incubación, se sacó la cámara Transwell y las células se fijaron con paraformaldehído al 4% y se tiñeron con violeta cristal al 0,1%. El tamaño de la cicatrización de la herida y el número de células de migración se calcularon para evaluar la capacidad de migración.
Determinación del ciclo celular
Después de la incubación durante la noche en placas de 6 pocillos, las células se trataron con hGC33 (Ab) libre, SFB libre, hGC33-null-NP o hGC33-SFB-NP durante 24 h, luego se recogieron y fijaron con etanol. Después de teñir con yoduro de propidio, se realizó una citometría de flujo y se analizó el ciclo celular con el modifit 3.0 (Verity Software House, Topsham, ME).
Western Blot
Para evaluar el estado de activación de la vía de la señal y la expresión de las moléculas diana, las células se incubaron durante la noche en placas de 6 pocillos y se aplicaron hGC33 libre, SFB libre, hGC33-null-NP o hGC33-SFB-NP durante 24 h. . Se recogieron las células de cada grupo de tratamiento y se extrajeron y midieron las proteínas. La concentración de proteína se midió y calibró con el kit de proteína BCA (Biosharp, Hefei, China). La proteína de las muestras se separó mediante electroforesis en gel de poliacrilamida con sulfato de alquilo doce, se transfirió a una membrana de PVDF y se selló con leche descremada. El primer anticuerpo (diluido 1:1000) se incubó durante la noche a 4 ° C y los segundos anticuerpos (1:2000) se incubaron durante 1 ha temperatura ambiente. Las bandas se visualizaron con sustratos ECL (Thermo Fisher Scientific Waltham, MA, EE. UU.), Y las imágenes se mostraron mediante un sistema de análisis de imágenes en gel y se utilizó β-actina como control.
Actividad antitumoral in vivo
Se determinó la inhibición de hGC33 libre, SFB libre, hGC33-null-NP, SFB-NP y hGC33-SFB-NP sobre el crecimiento de HCC in vivo. De acuerdo con las regulaciones y pautas sobre salud animal del comité de ética de la Universidad Tecnológica de Anhui, todos los experimentos se realizaron en ratones BALB / c en jaulas en una sala de control de temperatura (23 ± 2 ° C) con 12 h / 12 h de luz / ciclo oscuro. Una suspensión de 50 μl que contiene 5 × 10 6 Se inyectaron células HepG2 vivas por vía subcutánea en el abdomen derecho de ratones hembra BALB / c de 5 semanas de edad (20-22 g). Cuando el volumen del tumor alcanzó unos 50 mm 3 , los ratones se dividieron aleatoriamente en 6 grupos (10 ratones en cada grupo). Solución salina normal NS de control (200 mg / kg NP nula en 200 μL de PBS), hGC33-null-NP (hGC33-null-NP en 200 μL de PBS, equivalente a hgc33 =100 μg / kg / hora), hGC33 libre (hGC33 en 200 μL de PBS, 100 μg / kg / hora), SFB libre (dosis de SFB:8 mg / kg / hora), SFB-NP (dosis de SFB:8 mg / kg / hora) y hGC33-SFB-NP (equivalente a SFB =8 mg / kg / hora, hGC33 =100 μg / kg / hora) se inyectaron a través de la vena de la cola cada 2 días durante 10 veces. El peso y el tamaño del tumor de los ratones se midieron cada cuatro días. La fórmula de cálculo del volumen tumoral fue volumen =0,5 × L × W 2 , donde L y W representan la longitud y el ancho del tumor, respectivamente. Cuatro semanas después de la administración, los animales se anestesiaron con éter dietílico y se midieron el tamaño y el peso del tumor. Además, se extirparon el tumor, el corazón, el hígado, el riñón, el pulmón y el bazo, se fijaron con una solución de paraformaldehído al 4%, se incluyeron en parafina, se seccionaron y se tiñeron con hematoxilina y eosina para evaluar los cambios histológicos mediante microscopía digital.
Análisis estadístico
Los datos se presentan como la media ± desviación estándar (DE) y se evaluaron mediante análisis de varianza con SPSS 18.0. Se realizaron comparaciones estadísticas por pares utilizando una prueba t de Student de dos colas. Las diferencias se consideraron estadísticamente significativas para P <0.05.
Resultados
Caracterización de NP y liberación de fármacos In Vitro
Debido a que el diámetro y las propiedades de la superficie de las NP afectan la captación celular, la liberación de fármacos y la distribución de NP in vivo, caracterizamos las NP preparadas con los parámetros correspondientes. La morfología, el tamaño de partícula y la distribución del tamaño de partícula de los polímeros SFB-NP, hGC33-null-NP y hGC33-SFB-NP se resumen en la Fig. 1. La morfología de hGC33-SFB-NP observada por microscopio electrónico de transmisión (Fig. . 1a) muestra núcleos rígidos con bordes difusos, lo que indica que hGC33 está presente en la superficie de las NP. Los tamaños de partícula de SFB NP, hGC33-null-NP y hGC33-SFB-NP variaron de 100 a 150 nm y tenían una distribución típica de tamaño de partícula unimodal. El diámetro medio de hGC33-SFB-NP (120,2 ± 10,2 nm) fue ligeramente mayor que el de hGC33-null-NP, SFB-NP y nulo NP (Fig. 1a, Tabla 1). En la superficie de hGC33-SFB-NP y hGC33-null-NP estaban presentes películas esféricas delgadas con una sola superficie, lo que indicaba que el anticuerpo hGC33 estaba presente en la superficie de las NP. El tamaño aumentado para hGC33-SFB-NP y hGC33-null-NP confirmó la existencia de película de hGC33. La síntesis de hGC33-SFB-NP se confirmó con 1 H-NMR (Fig. 1b) antes de la preparación de NP. Los picos a 5,2 ppm y 1,58 ppm se asignaron a protones -CH3 del ácido láctico; el pico de 4,8 ppm se asignó a -OCH2- del ácido glicólico; y el pico de 3,6 a 3,8 ppm se asignó a los protones -CH2CH2O- de las unidades de repetición de PEG. Química de superficie de PEG- b -PLGA y Ab-PEG- b -Las NP de PLGA también se estudiaron mediante espectroscopia FTIR (Fig. 2c). En el espectro de PEG- b -Polímero PLGA, una banda fuerte a aproximadamente 1750 cm −1 se originó a partir del tramo de los grupos carbonilo (C =O) en la cadena PLGA. Una banda a 2880 cm −1 se debió al estiramiento de un grupo a –CH en la cadena PEG. Al mismo tiempo, apareció un pico a 2520 cm −1 que atribuimos al pico de estiramiento -SH del anticuerpo hGC33. El 1 Los resultados de H NMR y FTIR indicaron que el anticuerpo se injertó en la columna vertebral del PEG- b -Polímeros PLGA.
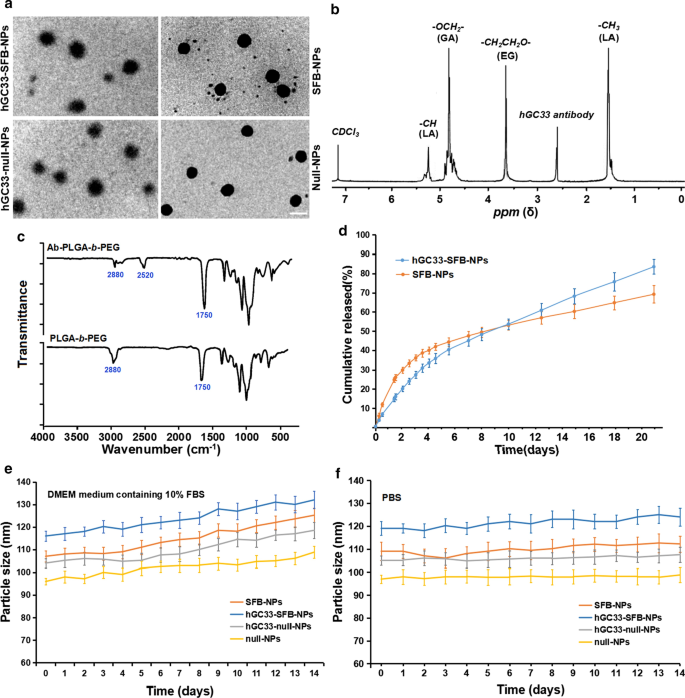
Caracterización de NP. un Caracterización TEM de NP, la barra de escala indica 100 nM; b el 1 Espectros de H RMN de hGC33-PLGA- b sintético -PEG en CDCl3; c Espectros FTIR de hGC33-PEG- b -PLGA y PEG- b - PLGA; d perfiles de liberación acumulativa de SFB-NP y hGC33-SFB-NP en PBS (pH =7,4) a 37 ° C; e cambios de tamaño de las NP incubadas en medio DMEM que contenía FBS al 10% durante 14 d; f cambios de tamaño de las NP incubadas en PBS durante 14 d. SFB, sorafenib; TEM, microscopía electrónica de transmisión; NP, nanopartículas; 1 H NMR, 1 Espectroscopia de resonancia magnética nuclear H; FTIR, espectroscopia infrarroja por transformada de Fourier
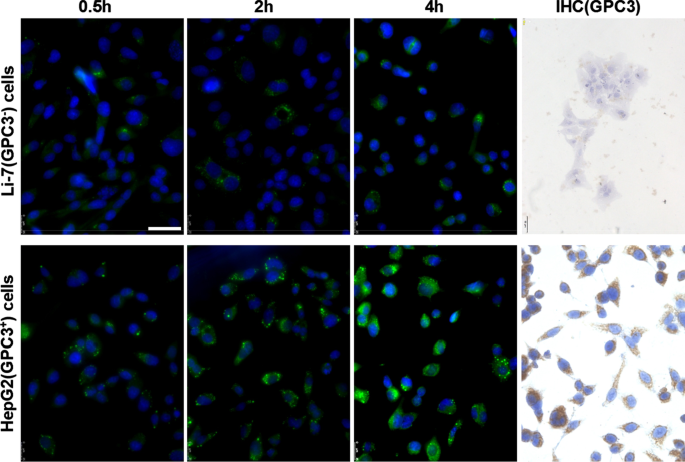
Expresión de GPC3 y captación de hGC33-cumarina6-NP en células Li-7 y HepG2. Las células, inoculadas en la placa de cultivo, se lavaron con PBS y se incubaron con 100 µg / ml de hGC33-cumarina6-NP en DMEM durante 2, 4 y 8 h. Los núcleos se tiñeron con DAPI y las células se fijaron y detectaron mediante microscopía de barrido láser confocal. No se detectó GPC3 en las células Li-7, pero se expresó en gran medida en las células HepG2 según se detectó mediante inmunocitoquímica. La barra de escala indica 50 μM
Curiosamente, las nanopartículas de hGC33-SFB-NP y SFB-NP en medio DMEM que contenía FBS al 10% (pH =7,4) tuvieron una liberación de fármaco rápida hasta aproximadamente 4 días, luego una liberación de fármaco relativamente lenta y estable; la liberación acumulada de SFB de hGC33-SFB-NP y SFB-NP durante 20 días fue de aproximadamente 77% y 65%, respectivamente (Fig. 1d). La diferencia puede deberse a la molécula hidrófila en la superficie del PEG- b -Matriz de PLGA, que puede acelerar la degradación de las nanopartículas aumentando la hidratación y promoviendo así la hidrólisis. Para determinar la estabilidad de los NP preparados, se colocaron hGC33-SFB-NP, hGC33-null NP, null-NP y SFB-NP en medio DMEM que contenía FBS al 10% (pH =7,4) y en PBS (pH =7,4). ). Los tamaños de los distintos NP se mantuvieron estables durante más de 2 semanas. Se liberó SFB de hGC33-SFB-NP de manera sostenida y estable durante 14 días, pero hubo un cambio modesto en el tamaño de las partículas en medio DMEM en comparación con el de FBS al 10% (Fig. 1e, f). La estabilidad de hGC33-SFB-NP sería apropiada para que SFB tenga una función biológica sostenida.
Un gran potencial zeta puede causar una fuerte interacción repulsiva electrostática entre NP y mantener la estabilidad del sistema de dispersión NP [23, 24]. Como se muestra en la Tabla 1, los potenciales zeta de hGC33-SFB-NP, hGC33-null-NP y SFB-NP son - 18,2 ± 2,2 mv, - 18,5 ± 1,8 mv y - 15,9 ± 2,1 mv, respectivamente, que pueden ser causada por las cargas negativas generadas por el grupo aldehído, el grupo carboxilo y el grupo fosfato en la glicoproteína hGC33 del anticuerpo. Los NP cargados negativamente pueden conducir a la alta estabilidad de la suspensión NP. Además, debido a que la superficie de las células está cargada negativamente en entornos fisiológicos normales, las NP preparadas repelen las células con baja carga y son menos tóxicas para los tejidos y las células. Además, la distribución de tamaño de nulo-NP y SFB-NP (índice de polidispersidad [PDI] 0,18 y 0,19, respectivamente) fue leve pero no significativamente menor que la de hGC33-null-NP (PDI =0,21) y hGC33-SFB- NP (PDI =0,23) Por lo tanto, el tamaño de las nanopartículas tiene una buena uniformidad. El tamaño de partícula, la distribución del tamaño de partícula y el potencial zeta de NP se muestran en la Tabla 1 de acuerdo con los resultados de la eficiencia de encapsulación de SFB y el contenido de carga. El anticuerpo GPC3 no conjugado hGC33 se eliminó mediante ultracentrifugación y se evaluó la eficacia de unión del anticuerpo hGC33 a las NP. El análisis de la proteína BCA mostró que la eficiencia de unión del anticuerpo hGC33 a las NP fue del 79,5% ± 2,9%.
hGC33-Coumarin 6-NP se dirige eficazmente a GPC3 + Células HepG2 de la línea celular HCC
Para averiguar si el anticuerpo hGC33 de las nanopartículas todavía tiene la capacidad de dirigirse específicamente a GPC3, usamos GPC3 + HepG2 y GPC3 - Células Li-7 como células diana y hGC33-cumarina 6-NP como nanopartículas trazadoras, y se incubaron las diferentes células durante 2 h, 4 hy 8 h. Las células se lavaron con PBS 3 veces y se hicieron reaccionar con DAPI para teñir el núcleo. Las células se fijaron y fotografiaron con un microscopio de fluorescencia Leica (DMi8, Alemania). Se encontró que la fluorescencia verde en las células HepG2 era significativamente mayor que la de las células Li-7 en el mismo tiempo de incubación (Fig.2), lo que indica que la cantidad de hGC33-cumarina 6-NP que entra en las células HepG2 fue significativamente mayor que la en celdas Li-7. Los resultados documentaron que el anticuerpo hGC33 en hGC33-Coumarin6-NP todavía tenía la capacidad de apuntar a GPC3 y mediar en la internalización de nanopartículas. La expresión de GPC3 en células HepG2 y células Li-7 se examinó con fluorescencia indirecta y tinción citoquímica. Los resultados mostraron que las células HepG2 sobreexpresaban GPC3, mientras que las células Li-7 no expresaban GPC3 (Fig. 2).
hGC33-Null-NP inhibe la proliferación de células HepG2
Para determinar si la NP modificada con hGC33 (hGC33-null-NP) podría inhibir el crecimiento de HCC, medimos la inhibición del crecimiento celular de la línea celular de HCC positiva para GPC3 HepG2 y la línea celular negativa para GPC3 Li-7. Encontramos que tanto hGC33-null-NP como hGC33 inhibieron el crecimiento de la línea celular de HCC positiva para GPC3 HepG2 después de 24 h de tratamiento, pero hGC33 tuvo un efecto inhibidor más significativo sobre las células HepG2 que hGC33-null-NP (Fig. 3a); por el contrario, hGC33-null-NP y hGC33 no afectaron al crecimiento de la línea celular Li-7 negativa para GPC3 (Fig. 3b). La evolución temporal representativa de hGC33-null-NP (equivalente a 0,1 µm de hgc33) y hGC33 (0,1 µm) en la proliferación de células HepG2 se ilustra en la Fig. 3c. Debido a que hGC33 es un péptido C-terminal que reconoce GPC3, hGC33 es superior a hGC33-null-NP en la inhibición de HepG2. Los resultados sugieren que la molécula hGC33 en hGC33-null-NP puede tener un efecto de bloqueo del espacio, lo que puede afectar la capacidad de unión de hGC33 a los epítopos.
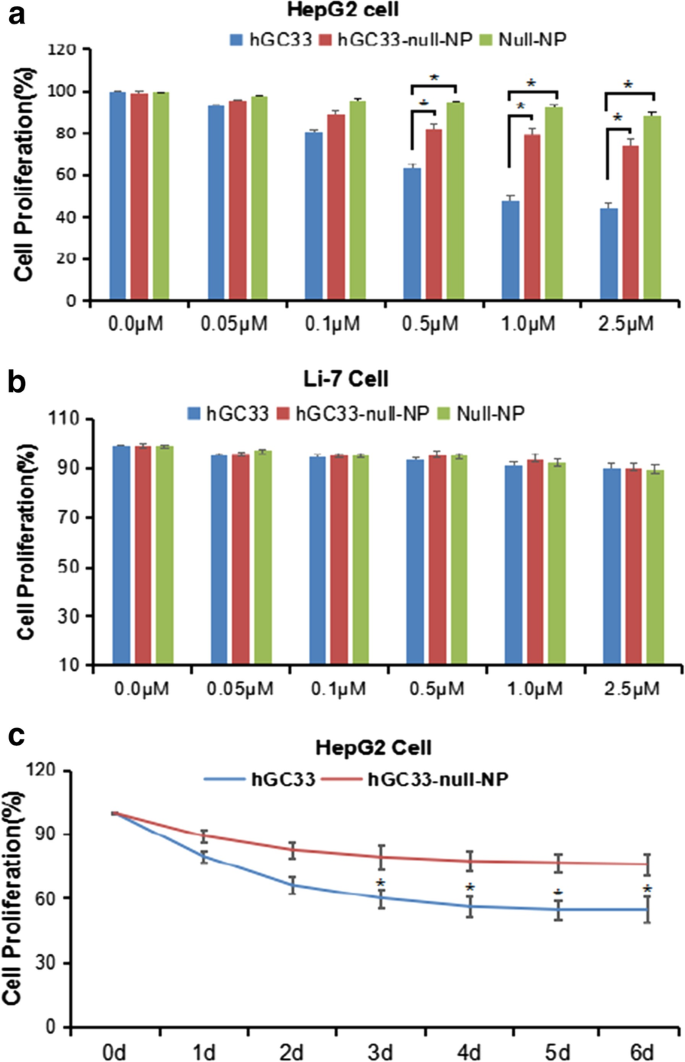
La proliferación de células HepG2 y Li-7 fue inhibida por hGC33, null-NP y hGC33-null-NP. un La prueba de proliferación celular se realizó en células HepG2 positivas para GPC3 tratadas con hGC33, nulo-NP o hGC33-nulo-NP; b Las pruebas de proliferación celular se realizaron en células Li-7 negativas para GPC3 tratadas con hGC33, nulo-NP y hGC33-nulo-NP. Las células se incubaron con hGC33 0-2,5 μM, nulo-NP o hGC33-nulo-NP durante 1 día. La proliferación de las células se midió con el método MTT y se estandarizó como células no tratadas. Todos los valores representan la media ± DE. En comparación con el grupo de control (0 μM) sin tratamiento con anticuerpos, * P <0,01; c los resultados representativos de la respuesta de HepG2 a hGC33, null-NP y hGC33-null-NP en el tratamiento con hGC33 (1.0 μM)
hGC33-Null-NP inhibe el ciclo celular de las células HepG2
Para comprender el mecanismo de la actividad molecular del anticuerpo hGC33 modificado en nanopartículas de hGC33-null-NP, estudiamos la progresión del ciclo celular de HepG2 tratada con hGC33-null-NP. En la línea celular HepG2, hGC33-null-NP y hGC33 aumentaron significativamente la proporción de células en G1 (Fig. 4a, b), lo que indica que tanto hGC33-null-NP como hGC33 podrían inducir la detención del ciclo celular en la fase G1. Además, hGC33-null-NP y hGC33 podrían regular significativamente la expresión de ciclinaD1 en células HepG2 (Fig. 4c, d).
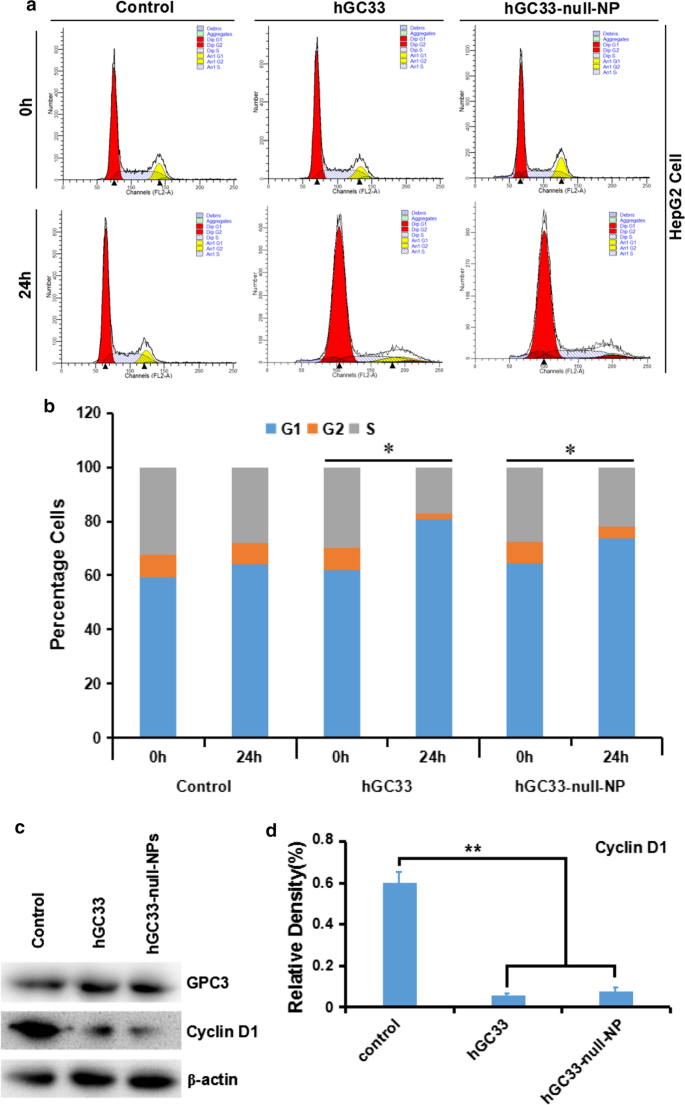
La detención del ciclo celular inducida por hGC33-null-NP y hGC33 en la fase G1 e inhibió la expresión de ciclinaD1 en células HepG2. un Diagrama representativo del ciclo celular de varios grupos tratados con hGC33-null-NP y hGC33. b Análisis del ciclo celular de varios grupos tratados con hGC33-null-NP y hGC33. Las células de HCC se incubaron con 0,5 µm de hGC33 o hGC33-null-NP (con la concentración molar de hGC33 como referencia). * P <0,05, la fase G1 de las células hGC33-null-NP o hGC33 se comparó con la de las células 0 h. c , d Después del tratamiento con hGC33-null-NP o hGC33, la ciclinaD1 se redujo significativamente en las células HepG2 en comparación con la del grupo de control
Activación de Wnt en celdas HepG2, Huh-7 y Li-7
Para comprender la activación de la señal clásica de Wnt / β-catenina en las células de HCC, primero detectamos la expresión del ligando de Wnt y la proteína de rizado del receptor (frizzled, FZD o Frz) en varias líneas celulares de HCC:HepG2 (GPC3 ++ ), Huh-7 (GPC + ) y Li-7 (GPC3 - ). Los resultados mostraron que el receptor Wnt3a y FZD de transducir la vía de la β-catenina se expresaron en todas las líneas celulares, especialmente en las células HepG2 y Huh-7 (Fig. 5).
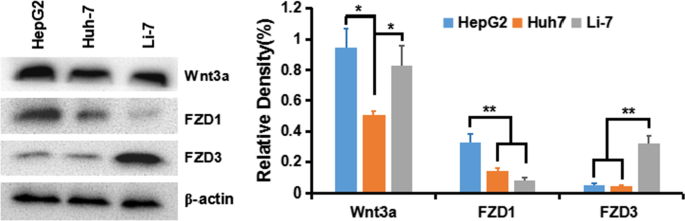
Expresión de Wnt3a, FZD1 y FZD3 en líneas celulares de HCC
hGC33-Null-NP inhibe la proliferación de células dependientes de la transducción de señales de Wnt inducida por Wnt3a
Algunos estudios han demostrado que la parte extracelular de GPC3 puede ser un correceptor de Wnt, que promueve la activación y transducción de la señal de Wnt / β-catenina. Por lo tanto, cuando HepG2 (GPC3 + ), Huh-7 (GPC3 + ) y Li-7 (GPC3 - ) cells were co-incubated with hGC33 and hGC33-null-NP, the activation of Wnt/β-catenin signal was blocked by hGC33 and hGC33-null-NP, and proliferation of HepG2 and Huh-7, but not Li-7, in Wnt3a-conditioned medium was inhibited. That proliferation of Li-7 cells most likely is due to the absence of GPC3 on the surface of Li-7 cells (Fig. 6). Our results indicate hGC33 and hGC33-null-NP nanoparticles specifically bind to GPC3 molecules on cell membrane. hGC33 and hGC33-null-NP partially neutralize the mitogenic activity of Wnt3a and inhibit the Wnt/β-catenin signaling pathway. However, the inhibition of proliferation by hGC33-null-NP nanoparticles was less than that of hGC33. Perhaps the spatial structure of the nanoparticles interferes with hGC33-null-NP and limits the function of the hGC33 molecule on the nanoparticles so they cannot completely block the interaction between GPC3 and Wnt3a.

GPC3-activated Wnt signal transduction in HCC. Fifty percent Wnt3a DMEM medium was added with anti-wnt3a antibody, hGC33, or hGC33-null-NP. HepG2 (GPC3 ++ ), Huh-7 (GPC3 + ) and Li-7 (GPC3 − ) cells were co-incubated for 48 h, and cell proliferation was measured by MTT assay. The data were expressed as mean ± SD (*P < 0.01)
hGC33-Null-NP Inhibits Wnt3a-Induced Signal Transduction in HepG2 and Huh-7 Cells
To understand the effect of hGC33-null-NP on Wnt/β-catenin signaling in HCC cells, we extracted the proteins of HepG2 and Huh-7 cells treated with hGC33 or hGC33-null-NP. The results of western blot showed that after hGC33-null-NP treatment, the levels of pYAP and pβ-catenin were increased, but the levels of YAP and β-catenin were decreased. Furthermore, the levels of cyclinD1, CD44, VEGF, and c-MYC in the hGC33-null-NP group were lower than those in the control group, but the level was less than with hGC33 treatment. Similar effects were observed in HepG2 and Huh-7 cells, as shown in Fig. 7.

Inhibition of Wnt3a-induced β-catenin signaling by hGC33-null-NP or hGC33. un Compared with the control group, Wnt/β-catenin signaling pathway in HepG2 and Huh-7 cells treated with hGC33-null-NP or hGC33 was inhibited, and the levels of β-catenin and YAP were decreased, while the increased phosphorylated β-catenin and phosphorylated YAP molecules were unstable, and degraded in cytoplasm. The decreased β-catenin was difficult to maintain in the nucleus and drive the expression of CyclinD1, CD44, VEGF, and c-MYC, which resulted in the decrease of cyclinD1, CD44, VEGF, and c-myc protein levels. b The mechanism pattern of Wnt/β-catenin signaling pathway inhibited by hGC33-null-NP or hGC33
hGC33-SFB-NP or hGC33 Attenuates HCC Cell Migration by Inhibiting Epithelial Mesenchymal Transition (EMT)
HCC is one of the deadliest cancers in the world, and its incidence is steadily increasing. Sorafenib is the only approved standard treatment for patients with advanced HCC, as it has been shown to improve the survival rate of these patients. However, clinical and preclinical observations suggest that sorafenib therapy has limited efficacy due to the rapid development of drug resistance. Therefore, elucidating the mechanism of escape resistance to sorafenib is a major emphasis in HCC research. In recent years, more and more attention has been paid to the role of epithelial mesenchymal transition (EMT) in the progress of HCC and the development of drug resistance. EMT refers to the transformation of epithelial to mesenchymal cells, which endows cells with the ability metastasize and invade, including acquisition of stem cell characteristics, reducing apoptosis and aging, promoting immunosuppression, and participating in the occurrence and development of cancer. The loss of E-cadherin expression is considered a key step in the carcinogenesis and EMT. EMT is a developmental multi-step molecular and cellular reprogramming process that cancer cells use to achieve aggression. This is mainly through down regulating the expression of E-cadherin, keratin, mucin, ZO-1 (tight junction protein); up regulating the expression of vimentin, alpha-smooth muscle actin (α-SMA), FN fibronectin, MMPs (degradation matrix), N-cadherin, snail, slug, twist, Rho, TGF-β, FGF, type I collagen, and type II collagen to achieve the invasion, metastasis, and anti-apoptosis of EMT characteristic tumor. The changes of these protein expressions mainly involve the activation of Wnt/β-catenin and Ras/Raf/MAPK signaling pathways.
Our experiments have shown that hGC33 antibody on the surface of NP vector can inhibit Wnt3a-induced β-catenin signal transduction, and then down regulate the expression of β—catenin, CD44, vascular endothelial growth factor (VEGF), cyclin D1, and c-MYC. Furthermore, hGC33-SFB-NP inhibits the activation of Ras/Raf/MAPK signal pathway and inhibits proliferation and apoptosis of HCC cells. hGC33 and SFB have a synergistic effect, inhibiting EMT and decreasing the migration of HCC cells (Fig. 8).

Effect of hGC33-SFB-NP on EMT inhibition. un Compared with the control group, the hGC33-SFB-NP treatment group had less cell migration. Photographs were taken under an optical microscope (magnification, × 200). The error value represents the standard deviation of three independent experiments. *Compared with the control group, p <0,01. b Compared with the control group, the EMT-related proteins snail, vimentin, and MMP-2 in HCCs treated with hGC33-SFB-NP decreased, whereas E-cadherin increased. c Molecular mechanism of EMT. EMT, epithelial–mesenchymal transition; MMP-2, matrix metalloproteinase-2; SFB, sorafenib; NP, nanoparticle
hGC33-SFB-NP Inhibits the Growth of Hepatocellular Carcinoma In Vivo and Improves the Survival Rate of Tumor-Bearing Mice
To evaluate the anti-tumor activity of hGC33-SFB-NP in vivo, HepG2 and Huh-7 cells were inoculated subcutaneously into the right abdomen and dorsal side of female BALB/c nude mice, respectively. When the tumor xenograft growth reached about 30 mm 3 , the mice were randomly divided into groups to further evaluate the inhibition of each group (hGC33-SFB-NP, hGC33-null-NP, SFB-NP, free hGC33, free SFB, and control group) HCC effect (n = 5 per group). It can be seen from Fig. 9a, b that hGC33-SFB-NP significantly slowed tumor growth in mice compared with the PBS control and other treatments. Compared with the PBS control, hGC33-null-NP, SFB-NP, free hGC33, and free SFB also had some inhibition of HCC, which is because free hGC33 and free SFB directly inhibit Wnt signal and Ras/Raf/MAPK, respectively. Such pathways can inhibit the proliferation of HCC cells to a certain extent. Although the nanoparticle-modified hGC33 (hGC33-null-NP) is connected to the nanosurface through chemical bonds, it did not affect hGC33′s targeting of GPC3 molecules and inhibition of Wnt activity. Nanoparticle-loaded SFB (SFB-NP), after being endocytosed by cells, was degraded to release SFB from the copolymer to inhibit the growth of HCC. In all, the inhibitory effect of hGC33-SFB-NP on HepG2 cell grafts was, as expected, more than on Huh-7 cell grafts, probably because HepG2 expresses GPC3 molecules.

The effect of hGC33-SFB-NP on xenotransplantation of HCC in nude mice and the changes of mice weight. Liver cancer cells were inoculated subcutaneously on the back of each nude mouse (n =10). After 10 days, the tumor bearing mice were treated with PBS (control), free hGC33, free SFB, hGC33-null-NP, SFB-NP, and hGC33-SFB-NP. Tumor size (a , b ) and body weight (c , d ) of mice were monitored at designated time points
The body weight of nude mice in each treatment group also was measured, as shown in Fig. 9c, d. The body weight of the control group decreased gradually. The weight of mice in free hGC33, free SFB, SFB-NP, and hGC33-null-NP treatment groups also decreased progressively and not significantly less than in the control group. However, the weight of nude mice bearing HepG2 and Huh-7 treated with hGC33-SFB-NP only slightly decreased, and the weight remained relatively stable during the treatment cycle. These results support that the novel hGC33-SFB-NP nanodrug has no significant toxicity in nude mice, and the SFB loaded on the nanocarrier and the surface modified hGC33 can produce additive or even synergistic anti-tumor effect.
Discussion
To examine the suitability of hGC33-SFB-NP for targeted HCC therapy, we tested the model conjugates for their ability to bind to human glypican-3 on HCC cells in vitro; to inhibit glypican-3-positive HCC cell proliferation, migration, and Wnt/β-catenin signal transduction; and inhibit HCC that overexpress glypican-3 in vivo.
To covalently bind GPC3-specific antibody hGC33 with mal-PEG-b -PLGA nanoparticles, we cross-linked the free sulfhydryl group in the Fc segment of hGC33 with maleimide functionalized PEG-b -PLGA (mal-PEG-b -PLGA) by forming stable thioether bonds. Conjugation is a prerequisite for targeting of GPC3-positive HCC. A series of experiments, including the changes of nanoparticle diameter and zeta potential detected by lens and the intracellular uptake of hGC33-SFB-NP, verified the targeting of hGC33-SFB-NP to HepG2 (GPC3 + ) células. These results indicated that the binding activity of antibody hGC33 was not altered by the conjugation.
We directly detected the phagocytic effect of GPC3 + HepG2 and GPC3 − Li-7 cells on PEG-b -PLGA NP surface-modified hGC33 by confocal microscopy. After HepG2 and Li-7 cells were co-cultured with hGC33-coumarin 6-NP, the green signal intensity in HepG2 cells was significantly higher than in Li-7 cells, indicating that there were more nanoparticles in the HepG2 cells. This finding is consistent with the hGC33 antibody modified on the surface of PEG-b -PLGA NP specifically binding to glypican-3 on the surface of HCC cells and being internalized. The efficiency of hGC33-modified NP internalization depends on the expression level of GPC3 antigen on the cell surface.
We used the standard MTT assay to measure the efficiency of inhibiting the proliferation of hepatoma cells. Both hGC33-null-NP and hGC33 inhibited the growth of the GPC3-positive HCC cell line HepG2, but hGC33-null-NP and hGC33 did not affect the proliferation of GPC3-negative Li-7 cells (Fig. 3b). At the animal level, hGC33-null-NP or hGC33 alone inhibited the growth of Huh-7 and HepG2 xenografts to a certain extent, while hGC33-SFB-NP caused growth arrest of Huh-7 and HepG2 hepatoma xenografts in mice. The finding that hGC33-null-NP significantly inhibited GPC3-positive hepatoma cells showed that the inhibitory effect of PEG-b -PLGA NP surface-modified hGC33 on HCC cell proliferation depends on the expression of GPC3 antigen on the cell surface.
GPC3 regulates many pathways in HCC pathogenesis, including Wnt and YAP signaling [25,26,27]. GPC3 interacts with Wnt ligand and may be a coreceptor for Wnt and facilitate Wnt/Frizzled binding for HCC growth [28, 29]. We further examined the effect of nanodrug surface-modified hGC33 on Wnt signaling pathway in hepatoma cells. Like free hGC33, nanodrug surface-modified hGC33 inhibited the proliferation of hepatoma cells not only by blocking Wnt-induced signal transduction in HepG2 and Huh-7 cells of expressing GPC3, but also by inhibiting Wnt3a-induced β-catenin and YAP signal transduction. Previous studies have shown that YAP expression is regulated by β-catenin at the transcriptional level of HCC [30, 31]. In this study, free hGC33 and nanodrug surface-modified hGC33 inhibited Wnt3a-induced YAP activity, indicating that Yap/TAZ released from β-catenin complex can also initiate classic Wnt signaling transduction [32, 2]. These results indicate that typical Wnt and YAP cross talk through a variety of mechanisms. Compared with hGC33-null-NP and hGC33, hGC33-SFB-NP had stronger anti-proliferation and anti-migration ability in vitro and in vivo. Thus, hGC33 not only determines the specificity of HCC cells, but also increases the inhibitory effect of SFB on the proliferation and migration of HCC cells by blocking the key signals related to tumor growth, such as Wnt/β-catenin and Wnt/YAP signaling pathway.
Conclusion
Antibody hGC33 to glypican-3, a membrane protein that is overexpressed on hepatocellular carcinoma cells, increased binding of sorafenib-loaded polyethylene glycol-b-PLGA polymer nanoparticles (hGC33-SFB-NP) to glypican-3 on the cancer cells. Administration of the antibody-modified nanoparticles synergistically inhibited Wnt-induced signal transduction and Ras/Raf/MAPK signaling pathway; hepatocellular carcinoma cells were arrested in G0/1 phase by down-regulation of cyclin D1 expression, thus attenuating cancer cell migration by inhibiting epithelial–mesenchymal transition. hGC33-SFB-NP inhibited the growth of liver cancer in vivo and improved the survival rate of tumor-bearing mice.
Disponibilidad de datos y material
Yes, all data have presented in the manuscript.
Abreviaturas
- AKT/PKB:
-
Protein kinase B
- c-MET:
-
HGFR:Hepatocyte growth factor receptor
- EE%:
-
Encapsulation efficiency %
- EMT:
-
Epithelial mesenchymal transition
- FTIR:
-
Espectroscopia infrarroja por transformada de Fourier
- 1 RMN H:
-
1 H-nuclear magnetic resonance spectroscopy
- HCC:
-
Hepatocellular carcinoma
- LC%:
-
Drug loading %
- MAPK:
-
Mitogen-activated protein kinases
- PDI:
-
Índice de polidispersidad
- PI3K:
-
Phosphoinositide 3-kinase
- pRAD51:
-
Phospho-RAD51
- SFB:
-
Sorafenib
- TEM:
-
Microscopía electrónica de transmisión
- YAP:
-
Yes-associated protein-1
Nanomateriales
- Nanopartículas para la terapia del cáncer:avances y desafíos actuales
- Preparación y propiedades magnéticas de nanopartículas de espinela de FeMn2O4 dopadas con cobalto
- Nanopartículas como bomba de eflujo e inhibidor de biopelícula para rejuvenecer el efecto bactericida de los antibióticos convencionales
- Efectos sinérgicos de nanopartículas de Ag / BiV1-xMoxO4 con actividad fotocatalítica mejorada
- Influencia de la rigidez elástica y la adherencia de la superficie en el rebote de nanopartículas
- Avances recientes en métodos sintéticos y aplicaciones de nanoestructuras de plata
- Toxicidad de nanopartículas de CoFe2O4 recubiertas de PEG con efecto de tratamiento de curcumina
- Efecto del agente peptizante ácido sobre la relación anatasa-rutilo y el rendimiento fotocatalítico de nanopartículas de TiO2
- Nanopartículas de oro de forma redonda:efecto del tamaño de partícula y la concentración sobre el crecimiento de la raíz de Arabidopsis thaliana
- Síntesis verde de nanopartículas de metal y óxido de metal y su efecto sobre el alga unicelular Chlamydomonas reinhardtii
- Sobreenfriamiento de agua controlado por nanopartículas y ultrasonido