


El microARN-301 inhibido restringe la angiogénesis y el crecimiento celular en el carcinoma de células escamosas de esófago al elevar el PTEN
Resumen
Objetivo
El carcinoma de células escamosas de esófago (ESCC) se caracteriza por metástasis precoz y diagnóstico tardío. Se sabe que el microARN-301 (miR-301) participa en diversos cánceres. Sin embargo, los efectos de miR-301 en ESCC permanecen inexplorados. Por lo tanto, nuestro objetivo es explorar el papel de miR-301 en la progresión de ESCC.
Métodos
Se evaluó la expresión de miR-301 y el homólogo de fosfatasa y tensina (PTEN) en tejidos y líneas celulares de ESCC. A continuación, las células seleccionadas se trataron con oligonucleótido y plásmido miR-301 o PTEN alterados y, a continuación, se evaluaron la capacidad de formación de colonias, la viabilidad celular, la migración, la invasión, la distribución del ciclo celular y la apoptosis de las células ESCC. Además, también se evaluaron el crecimiento tumoral y la densidad de microvasos (MVD), y se afirmó la relación de orientación entre miR-301 y PTEN.
Resultados
MiR-301 se reguló positivamente y PTEN se reguló negativamente en los tejidos y células de ESCC. Se seleccionaron células KYSE30 y células Eca109 para ensayos funcionales. En las células KYSE30, el miR-301 inhibido o el PTEN sobreexpresado suprimieron los comportamientos malignos celulares, y el PTEN silenciado eliminó el impacto de la inhibición de miR-301 en la progresión de la ESCC. En las células Eca109, la sobreexpresión de miR-301 o la inhibición de PTEN promovieron comportamientos malignos celulares, y la sobreexpresión de PTEN revirtió los efectos de la elevación de miR-301 en la progresión de ESCC. El ensayo in vivo reveló que la inhibición de miR-301 o la sobreexpresión de PTEN reprimió el crecimiento tumoral de ESCC y MVD, y la elevación de miR-301 o la reducción de PTEN tuvieron efectos contrarios. Además, PTEN fue el objetivo del miR-301.
Conclusión
En conjunto, los resultados de nuestro estudio revelaron que miR-301 afectaba el crecimiento celular, la metástasis y la angiogénesis mediante la regulación de la expresión de PTEN en ESCC.
Introducción
El cáncer de esófago (CE), el octavo cáncer más común en todo el mundo, es una neoplasia maligna crítica con alta mortalidad y mal pronóstico [1]. El carcinoma de células escamosas de esófago (ESCC), que representa alrededor del 90% del total de casos de CE, es la forma principal de CE en China [2]. Múltiples causas, como el bajo nivel socioeconómico, el tabaquismo, el consumo de alcohol, la ingesta nutricional deficiente, los alimentos ricos en nitrosaminas o contaminados con micotoxinas, conducen a la aparición de ESCC [3]. A pesar de los resultados clínicos promovidos, así como la administración, todavía hay un mal pronóstico entre los pacientes con ESCC, lo que acompaña a una tasa de supervivencia a 5 años del 15-25% [4]. Por lo tanto, es crucial confirmar oncogenes o genes represores de tumores que podrían funcionar como biomarcadores en el desarrollo de ESCC para proporcionar métodos terapéuticos más efectivos para pacientes con ESCC.
Los microARN (miARN) son pequeños ARN no codificantes que desempeñan un papel esencial en la modulación de la expresión génica [5] y se aclara que tienen la capacidad de influir en la progresión tumoral mediante la regulación de la estabilidad del ARNm y la capacidad de los ARNm [6]. Se ha encontrado que una cantidad de miRNAs como miR-4324 [7], miR-889-3p [8] y miR-9 [9] están asociados con el proceso de ESCC. MiR-301 es un miembro de los miARN que está formado por la unidad de transcripción fam33a situada en 17q22-23 en el genoma humano. La sobreexpresión de miR-301 se ha identificado previamente, lo que refleja que está implicado en enfermedades humanas [6, 10]. Sin embargo, los mecanismos de función de miR-301 no se han descubierto en ESCC. Además, se ha afirmado que el homólogo de fosfatasa y tensina (PTEN) se altera con frecuencia en los tumores y es el objetivo de las mutaciones de la línea germinal en pacientes con cáncer, que desempeña un papel inhibidor de los tumores [11]. Se ha validado que la desregulación de PTEN se correlaciona con el desarrollo de ESCC [12]. Curiosamente, una investigación reciente ha revelado que el miR-301 se dirige a PTEN en el cáncer de pulmón de células no pequeñas [13]. Sin embargo, esta relación de focalización entre miR-301 y PTEN en el desarrollo de ESCC aún no se ha revelado. Nuestra investigación se centró en los efectos de miR-301 y PTEN en la progresión de ESCC, que siguen siendo en gran parte desconocidos y son una novedad. Inferimos que miR-301 puede influir en la angiogénesis y el crecimiento celular en ESCC modulando la expresión de PTEN.
Materiales y métodos
Declaración de ética
Se obtuvieron consentimientos informados por escrito de todos los pacientes antes del estudio. Los protocolos de este estudio fueron aprobados por el Comité de Ética del Segundo Hospital de la Universidad de Jilin y se basaron en los principios éticos para la investigación médica con seres humanos de la Declaración de Helsinki. Los experimentos con animales fueron estrictamente consistentes con la Guía para el manejo y uso de animales de laboratorio emitida por los Institutos Nacionales de Salud. El protocolo de experimentos con animales fue aprobado por el Comité Institucional de Uso y Cuidado de Animales del Segundo Hospital de la Universidad de Jilin.
Temas de estudio
Se recolectaron ciento diez muestras de tejidos ESCC y tejidos normales adyacentes (> 5 cm del tumor) de pacientes con ESCC (78 hombres y 32 mujeres) que habían aceptado la esofagectomía en el departamento de cirugía torácica del Segundo Hospital de la Universidad de Jilin desde enero. 2015 a diciembre de 2017. Entre los 110 pacientes, hubo 84 casos> 60 años y 26 casos ≤ 60 años; tamaño del tumor:65 casos ≥ 5 cm y 45 casos <5 cm; 71 casos sin metástasis ganglionares (LNM) y 39 casos con LNM; la etapa de tumor, ganglios y metástasis (TNM):60 casos estaban en la etapa I + II y 50 casos en la etapa III; localización del tumor:13 casos fueron ESCC superior y 97 casos fueron ESCC medio-inferior. Todos los pacientes fueron diagnosticados con ESCC y no habían aceptado radioterapia o quimioterapia antes. Los tumores fueron extirpados por completo y el margen quirúrgico negativo ha sido confirmado por patología. De acuerdo con los criterios de estadificación de ESCC que propuso la Unión para el Control Internacional del Cáncer (UICC) en 2009 [14], la estadificación patológica posoperatoria de los pacientes se identificó como la etapa pT1-4N1-2 (I-IIIb). No hubo complicaciones significativas en los pacientes después de la cirugía y se excluyeron las muertes perioperatorias.
Reacción en cadena cuantitativa de la polimerasa con transcripción inversa (RT-qPCR)
Los ARN totales en tejidos y células se extrajeron usando kits Trizol (Invitrogen Inc., Carlsbad, CA, EE. UU.). Se midieron la concentración y la calidad del ARN. Los cebadores de ARN (Tabla 1) fueron diseñados y sintetizados por TaKaRa Biotechnology Co., Ltd. (Liaoning China). El ARN se transcribió a la inversa en ADNc según las instrucciones del kit de reactivos Takara PrimeScript ™ RT con g DNA Eraser (Takara). Realizamos qPCR en el Light Cycler 480II (Roche) con la mezcla maestra de PCR Power PCR SYBR green (Takara). Se usó U6 como control de carga de miR-301 y β-actina como referencia interna de PTEN. Los datos se analizaron usando 2 - △△ Ct método [15].
Análisis de Western Blot
Se utilizó tampón de lisis RIPA (Beyotime Institute of Biotechnology, Shanghai, China) para extraer la proteína total en células y tejidos, y la proteína se cuantificó mediante un kit de ensayo de proteínas BCA (Beyotime). Se midió la concentración de proteína de cada muestra y se realizó electroforesis en gel de poliacrilamida-dodecilsulfato de sodio al 10%. Las muestras se transfirieron a las membranas de nitrocelulosa, que luego se bloquearon con leche desnatada en polvo al 5% a 4 ° C durante la noche. Posteriormente, las membranas se suplementaron con anticuerpos primarios PTEN y β-actina (ambos 1:500 y de Santa Cruz Biotechnology Inc, CA, EE. UU.) Para incubación durante la noche, luego se agregaron con los respectivos anticuerpos secundarios y se incubaron durante 1 h. Después de sumergirlas en el reactivo quimioluminiscente mejorado (Pierce Chemical Inc., Dallas, TX, EE. UU.) Durante 1 minuto, las membranas se expusieron en un ambiente oscuro y se revelaron usando el mini generador de imágenes de quimioluminiscencia LAS4000. Los valores de gris se evaluaron mediante un software de sistema de imágenes con β-actina como control; por tanto, se obtuvo la proteína relativa final expresada. Las bandas de proteínas se analizaron con el software ImageJ2x.
Ensayo génico indicador de luciferasa dual
Se predijo que la secuencia de la región 3 'sin traducir (UTR) de PTEN interactuaría con miR-301, o se sintetizó una secuencia mutada dentro de los sitios diana predichos y se insertó en los sitios XbaI y FseI del vector indicador de luciferasa de control pGL3 (Promega, WI, EE. UU.). Luego, se produjeron los vectores pGL3-PTEN-wt y pGL3-PTEN-mut. Los plásmidos informadores de luciferasa wt y mut identificados correctamente con miR-301 imita y imita NC se cotransfectaron en células KYSE30 y Eca109 durante 48 h. Posteriormente, las células se lisaron y las actividades de luciferasa se determinaron, respectivamente, mediante kits de detección de luciferasa (Promega).
Cultivo celular, agrupación y transfección
Las líneas celulares ESCC (KYSE-150, KYSE-30, Eca109 y KYSE-70) se obtuvieron del Instituto de Bioquímica y Biología Celular de Shanghai, Academia de Ciencias de China (Shanghai, China), y se adquirieron las células epiteliales del esófago humano (HEEC) de Mingzhou Biotechnology Co., Ltd. (Zhejiang, China). Las células se cultivaron en medio RPMI 1640 (Invitrogen) suplementado con suero bovino fetal al 10% (FBS, Life Technologies, EE. UU.), 100 unidades / ml de penicilina G sódica (Sigma) y 100 μg / ml de sulfato de estreptomicina (Sigma). La expresión de MiR-301 y la expresión de ARNm de PTEN en cada línea celular se midieron mediante RT-qPCR, y se eligió la línea celular con la expresión relativa más alta y más baja para los experimentos celulares posteriores.
Las células KYSE-30 se separaron en 7 grupos y, respectivamente, se trataron con inhibidor de miR-301, control negativo del inhibidor (NC), pcDNA-PTEN (denominado sobreexpresado (oe) -PTEN), pcDNA-NC (denominado oe-NC), Inhibidor de miR-301 + ARN interferente pequeño (si) -PTEN o inhibidor de miR-301 + si-NC. Las células Eca109 se separaron también en 7 grupos y se trataron de forma solidaria con miR-301 mimic, mimic NC, si-PTEN, si-NC, miR-301 mimic + oe-PTEN, miR-301 mimic + oe-NC. el inhibidor NC, el inhibidor de miR-301, el imitador de miR-301, el imitador de NC, si-NC y si-PTEN se adquirieron de GenePharma Ltd., Company (Shanghai, China); pcDNA-PTEN NC y pcDNA-PTEN se obtuvieron de (Shanghai Sangon Bio-technology Corporation (Shanghai, China)). Las células se transfectaron transitoriamente en células ESCC por lipofectamine 2000 (Invitrogen) cuando la confluencia celular alcanzó el 60%.
Ensayo del kit de recuento celular (CCK-8)
Las células se sembraron en una placa de 96 pocillos (1 × 10 3 células / pocillo) y se incubaron durante diferentes períodos de tiempo. Después de incubar durante 24 h, 48 h, 72 hy 96 h, cada pocillo se suplementó con 10 μL de solución CCK-8 (5 mg / mL), y luego, las células de cada grupo se incubaron a 37 ° C sin luz. exposición durante 2 h. Los valores de densidad óptica (DO) a 450 nm se analizaron con un lector de microplacas (Bio-Rad Laboratories, Hercules, CA, EE. UU.).
Ensayo de formación de colonias
Las células se sembraron a 500 células / pocillo en placas de 6 pocillos después de la transfección y se cultivaron durante 14 d. Las colonias se fijaron con metanol, se tiñeron con violeta cristal al 0,5% y se contaron bajo el microscopio invertido.
Ensayo Transwell
Celdas (5 × 10 3 ) incubados en medio RPMI 1640 se sembraron en cámaras apicales de dispositivos Transwell con membrana no recubierta o recubierta con matrigel (Corning, NY, EE. UU.). Después de 24 h, se retiraron las células de las cámaras apicales, mientras que las células que quedaban en la parte inferior se fijaron y luego se tiñeron con violeta cristal al 0,1%. Se utilizó un microscopio (Olympus Corporation, Tokio, Japón) para contar 3 campos aleatorios para calcular el número de células.
Citometría de flujo
El ciclo celular y la apoptosis se evaluaron mediante citometría de flujo. Se incubaron isotiocianato de fluoresceína-anexina V (10 µL) y yoduro de propidio (PI; 5 µL, Sigma) con las células (5 × 10 5 células / pocillo) en la oscuridad a 4 ° C durante 30 min. El porcentaje de células apoptóticas se calculó utilizando un citómetro de flujo (BD Biosciences, CA, EE. UU.) Con el software FlowJo versión 10 (FlowJo LLC, OR, EE. UU.).
Para evaluar el ciclo celular, las células (5 × 10 5 células / pocillo) se fijaron con etanol al 75% durante la noche a 4ºC y se tiñeron con 5 \ mu l de PI / ribonucleasa A (Sigma) a 4ºC durante 30 min en la oscuridad. Los datos se analizaron con un citómetro de flujo (BD Biosciences). Las señales de fluorescencia (14.000) de cada muestra se recopilaron y calcularon utilizando el software ModFit LT versión 3.2 (Verity Software House, Inc., ME, EE. UU.).
Tumorigénesis subcutánea en ratones desnudos
Se obtuvieron 42 ratones desnudos hembra BALB / c-nu (de 4 semanas de edad, con un peso de 16-24 g) del Centro de Animales Experimentales de la Universidad de Jilin (Changchun, China). Los ratones desnudos se separaron en 14 grupos ( n =3). A los ratones desnudos de siete grupos se les inyectaron, respectivamente, células KYSE-30 de acuerdo con el agrupamiento de células, y a los ratones desnudos de los siete grupos restantes se les inyectaron por separado células Eca109 basándose en el agrupamiento. La concentración de células KYSE-30 y Eca109 transfectadas se ajustó a 5 x 10 6 células / 100 μL. Los ratones desnudos se fijaron y se inyectaron subcutáneamente con las células ESCC correspondientes en condiciones estériles. Se midieron semanalmente la mayor longitud (L) y ancho (W) de los tumores y el volumen del tumor ( V ) =1/2 × L × W 2 . Los ratones desnudos fueron sacrificados en el 5 th semana de inyección con los tumores resecados, y los tumores fueron pesados y fotografiados. La tasa de formación de tumores se calculó como el número de ratones con tumor subcutáneo / número total de ratones desnudos inyectados en el grupo x 100%. El tiempo de inyección se tomó en abscisas y el tamaño del tumor en ordenadas; por lo tanto, se trazó un gráfico de la curva de crecimiento del tumor.
Tinción inmunohistoquímica
Los tejidos tumorales de ratones desnudos se fijaron con formaldehído al 10%, se incluyeron en parafina y se seccionaron en 4 µm. A continuación, las secciones se secaron a 60 ° C durante 2 h, se desparafinaron con xileno, se deshidrataron con etanol en gradiente y se incubaron con 50 μL de H 2 al 3%. O 2 durante 10 min. Posteriormente, las secciones se empaparon en una solución tampón de ácido cítrico 0,01 M, se hirvieron a 95 ° C durante 20 min, se bloquearon con una solución de trabajo de suero de cabra normal a 37 ° C durante 10 min y se agregaron CD 34 (1:100, Santa Cruz) a 4 ° C durante la noche. Posteriormente, las secciones se suplementaron con polímero IgG de cabra anti-conejo / ratón marcado con HRP (ZSGB-Bio, Beijing, China), se contratiñeron con hematoxilina, se deshidrataron y permeabilizaron y luego se sellaron con bálsamo neutro. Se usó PBS para reemplazar los anticuerpos primarios como NC. Medición de la densidad de microvasos (MVD):las secciones se observaron bajo un microscopio de bajo aumento. Se tomaron como microvasos una célula endotelial o un grupo de células endoteliales teñidas de amarillo parduzco y se distinguieron significativamente con las células tumorales circundantes, y los tejidos conectivos. La estructura de la rama también se tomó como un vaso si estaba desconectada, mientras que se excluyeron los vasos con tamaño de lumen> 8 eritrocitos, capa muscular o lumen más grueso. Se registró el número de microvasos de 3 campos visuales altos y el número promedio fue MVD de cada caso.
Análisis estadístico
Todos los análisis estadísticos se llevaron a cabo utilizando el software SPSS versión SPSS 21.0 (IBM Corp. Armonk, NY, EE. UU.) Y se presentaron con Graphpad Prism Software 6.0. Los datos se expresaron como media ± desviación estándar. Las diferencias entre dos grupos independientes se probaron con la prueba t de Student. Se realizó un ANOVA de una vía para comparar tres o más grupos. P valor <0.05 fue indicativo de una diferencia estadísticamente significativa.
Resultados
MiR-301 tiene mucha expresión, mientras que PTEN se expresa mal en tejidos y células ESCC
La expresión de MiR-301 y PTEN en tejidos ESCC y tejidos normales adyacentes se evaluó mediante RT-qPCR y análisis de transferencia Western para revelar sus funciones en ESCC, y se encontró que (Fig. 1a-c) miR-301 estaba regulado al alza, mientras que PTEN fue regulada negativamente en los tejidos ESCC. Los pacientes se dividieron en grupos de expresión baja y alta según el valor mediano de la expresión de miR-301 o PTEN para analizar la correlación entre la expresión de miR-301 o PTEN y las características clínico-patológicas de los pacientes con ESCC. Los resultados indicaron que la expresión de miR-301 / PTEN no se relacionó con la edad, el sexo, el tamaño del tumor, la ubicación y la diferenciación, mientras que se correlacionó con el estadio TNM y LNM de los pacientes con ESCC (Tabla 2).
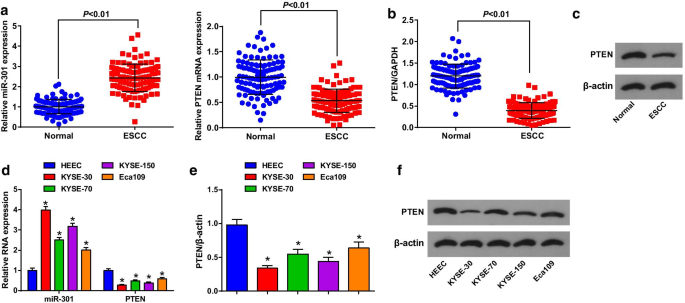
MiR-301 se expresa en gran medida, mientras que PTEN se expresa escasamente en tejidos y células de ESCC. un Expresión de miR-301 y expresión de ARNm de PTEN en el tejido ESCC detectada usando RT-qPCR; b expresión de proteínas de PTEN en el tejido de ESCC detectada mediante análisis de transferencia Western; c bandas de proteínas de PTEN en el tejido ESCC en análisis de transferencia Western; d expresión de miR-301 y expresión de ARNm de PTEN en la línea celular ESCC detectada usando RT-qPCR; e expresión de proteína de PTEN en la línea celular ESCC detectada usando análisis de transferencia Western; f bandas de proteínas de PTEN en análisis de transferencia Western. * P <0,05 frente a HEEC. Los datos se expresaron como media ± desviación estándar y se realizó la prueba t para las comparaciones entre dos grupos
Luego, se determinó la expresión de miR-301 y PTEN en 4 líneas celulares ESCC y HEECs usando RT-qPCR y análisis de transferencia Western. Encontramos que (Fig. 1d-f) miR-301 estaba regulado positivamente y PTEN regulado negativamente en las líneas celulares ESCC, entre las cuales KYSE-30 tenía la expresión más alta de miR-301 y la expresión PTEN más baja, mientras que Eca109 tenía la tendencia contraria. Por lo tanto, la línea celular KYSE-30 se trató con miR-301 regulado negativamente / PTEN sobreexpresado y la línea celular Eca109 se trató con miR-301 sobreexpresado / PTEN silenciado en los experimentos celulares.
PTEN está dirigido por miR-301
Un software bioinformático (http://www.microrna.org/) predijo que PTEN era el gen diana de miR-301 (Fig. 2a). Además, se confirmó mediante el ensayo del gen indicador de luciferasa dual que la actividad de la luciferasa se redujo significativamente en las células ESCC cotransfectadas con el vector PTEN-wt y el imitador de miR-301 en comparación con las cotransfectadas con el vector PTEN-mut y el imitador de miR-301, lo que implica que miR-301 podría unirse particularmente a PTEN (Fig. 2b, c).
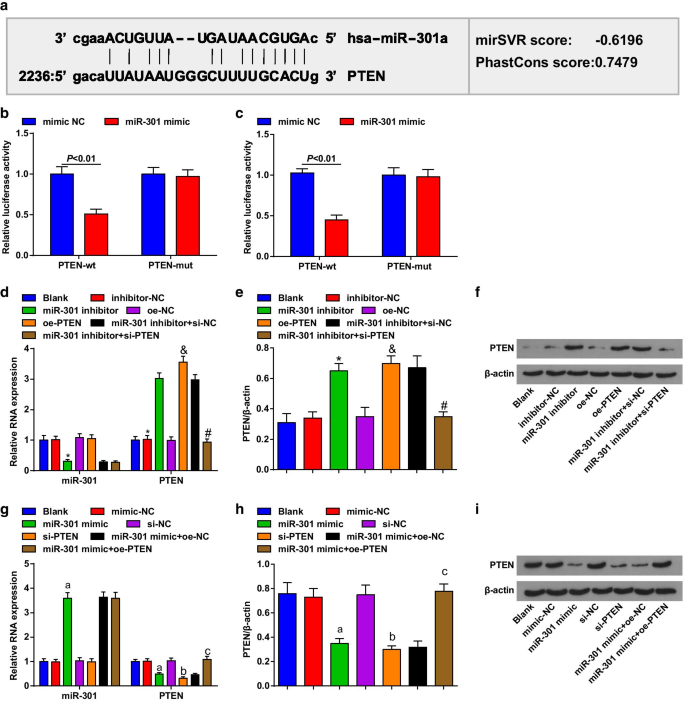
PTEN es el gen diana de miR-301. un Los sitios de unión de miR-301 y PTEN se predijeron mediante software de predicción en línea; b la relación diana entre miR-301 y PTEN en células KYSE-30 se evaluó mediante un ensayo de gen indicador de luciferasa dual; c la relación diana entre miR-301 y PTEN en células Eca109 se evaluó mediante un ensayo de gen indicador de luciferasa dual; d expresión de miR-301 y expresión de ARNm de PTEN en células KYSE-30 detectadas usando RT-qPCR después de regulación negativa de miR-310 o regulación positiva de PTEN; e expresión de proteína de PTEN en células KYSE-30 detectada usando análisis de transferencia Western después de regulación negativa de miR-310 o regulación positiva de PTEN; f bandas de proteínas de PTEN en células KYSE-30 en análisis de transferencia Western después de regulación negativa de miR-310 o regulación positiva de PTEN; g expresión de miR-301 y expresión de ARNm de PTEN en células Eca109 detectadas usando RT-qPCR después de regulación positiva de miR-310 o regulación negativa de PTEN; h expresión de proteína de PTEN en células Eca109 detectada usando análisis de transferencia Western después de regulación positiva de miR-310 o regulación negativa de PTEN; yo bandas de proteína de PTEN en células Eca109 en análisis de transferencia Western después de regulación positiva de miR-310 o regulación negativa de PTEN. * P <0,05 frente al grupo inhibidor-NC, & P <0.05 en comparación con el grupo oe-NC, # P <0,05 frente al grupo inhibidor de miR-301 + si-NC, a P <0.05 versus el grupo de imitación-NC, b P <0.05 versus el grupo si-NC, c P <0,05 frente al grupo mimético de miR-301 + oe-NC; N =3. Los datos se expresaron como media ± desviación estándar y se realizó la prueba t para las comparaciones entre dos grupos. Se utilizó ANOVA para comparaciones entre múltiples grupos
Se utilizaron RT-qPCR y análisis de transferencia Western para evaluar la expresión de miR-301 y PTEN en células transfectadas, y se encontró que en las células KYSE-30 (Fig. 2d-f), las células tratadas con el inhibidor de miR-301 regulaban negativamente miR-301 , mientras que PTEN regulaba al alza; las células tratadas con pcDNA-PTEN (oe-PTEN) elevaron la expresión de PTEN y si-PTEN revirtieron el efecto del inhibidor de miR-301 sobre la expresión de PTEN. En las células Eca109 (Fig. 2g-i), las células tratadas con miR-301 imitan a miR-301 regulado al alza, mientras que al PTEN regulado a la baja; las células tratadas con si-PTEN disminuyeron la expresión de PTEN, y pcDNA-PTEN (oe-PTEN) invirtió el papel inhibidor del imitador de miR-301 en la expresión de PTEN. Estos datos sugirieron que el miR-301 apuntó a PTEN.
El miR-301 inhibido o PTEN sobreexpresado restringe la viabilidad de las células ESCC; MiR-301 elevado o PTEN reducido promueve la viabilidad de las células ESCC
La viabilidad celular de las células ESCC se evaluó usando la formación de colonias y ensayos CCK-8. Los resultados revelaron que en la línea celular KYSE-30 (Fig. 3a-c), la transfección del inhibidor de miR-301 o oe-PTEN reprimió la capacidad de formación de colonias y la viabilidad celular; la transfección de PTEN silenciado eliminó el impacto del inhibidor de miR-301 en la viabilidad de las células ESCC; en la línea celular Eca109 (Fig. 3d-f), la transfección del imitador de miR-301 o la capacidad de formación de colonias promovida por si-PTEN y la viabilidad celular; La sobreexpresión de PTEN revirtió el papel promotor de la elevación de miR-301 en la capacidad de formación de colonias y la viabilidad de las células Eca109. Estos resultados sugirieron que la eliminación de miR-301 o la sobreexpresión de PTEN reprimieron la viabilidad de las células ESCC, que fueron promovidas por la elevación de miR-301 o la inhibición de PTEN.
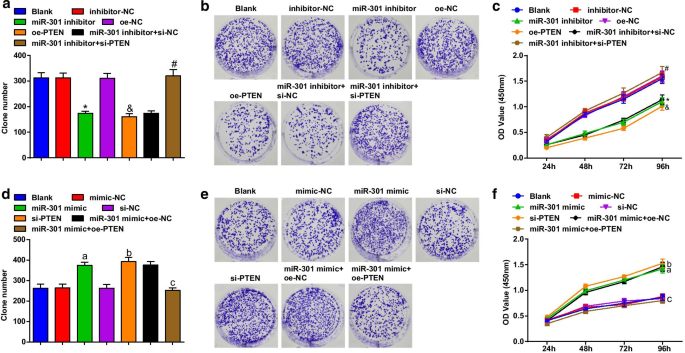
El miR-301 inhibido o PTEN sobreexpresado restringe la viabilidad de las células ESCC; miR-301 elevado o PTEN reducido promueve la viabilidad de las células ESCC. un Número de colonias en las células KYSE-30 después de la transfección detectadas usando un ensayo de formación de colonias después de la regulación a la baja de miR-310 o la regulación al alza de PTEN; b capacidad de formación de colonias de las células KYSE-30 después de la transfección detectada usando un ensayo de formación de colonias después de la regulación a la baja de miR-310 o la regulación al alza de PTEN; c viabilidad de las células KYSE-30 después de la transfección detectada usando el ensayo CCK-8 después de la regulación a la baja de miR-310 o la regulación al alza de PTEN; d número de colonias en células Eca109 después de la transfección detectadas usando un ensayo de formación de colonias después de la regulación positiva de miR-310 o regulación negativa de PTEN; e capacidad de formación de colonias de las células Eca109 después de la transfección detectada usando un ensayo de formación de colonias después de la regulación por aumento de miR-310 o regulación por disminución de PTEN; f viabilidad de las células Eca109 después de la transfección detectada usando el ensayo CCK-8 después de la regulación positiva de miR-310 o regulación negativa de PTEN; * P <0,05 frente al grupo inhibidor-NC; & P <0,05 frente al grupo oe-NC; # P <0,05 frente al grupo inhibidor de miR-301 + si-NC; a P <0,05 frente al grupo imitador-NC; b P <0,05 frente al grupo si-NC; c P <0.05 versus el grupo miR-301 mimic + oe-NC, N =3. Los datos se expresaron como media ± desviación estándar y se utilizó ANOVA para las comparaciones entre múltiples grupos
MiR-301 inhibido o PTEN sobreexpresado representa la migración e invasión de células ESCC; MiR-301 elevado o PTEN reducido induce la migración e invasión de células ESCC
Las capacidades de migración e invasión de las células ESCC se evaluaron utilizando el ensayo Transwell. Los resultados sugirieron que en la línea celular KYSE-30 (Fig. 4a, b), la migración celular y la capacidad de invasión estaban restringidas por la inhibición de miR-301 o la sobreexpresión de PTEN; el papel supresor del inhibidor de miR-301 en la migración celular y la capacidad de invasión fue revertido por si-PTEN. En la línea celular Eca109 (Fig. 4c, d), las capacidades de invasión y migración celular fueron promovidas después de la transfección del mimético de miR-301 o si-PTEN; El PTEN sobreexpresado revirtió el impacto del imitador de miR-301 en la migración celular y las capacidades de invasión. Los hallazgos anteriores implicaron que la migración e invasión de las células ESCC fueron inhibidas por la represión de miR-301 o la elevación de PTEN, mientras que fueron facilitadas por la regulación positiva de miR-301 o la regulación negativa de PTEN.
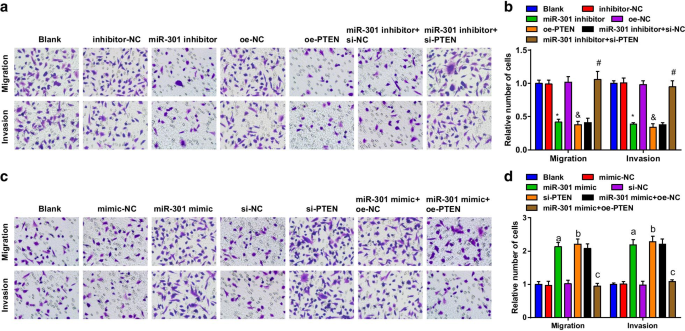
El miR-301 inhibido o PTEN sobreexpresado reprime la migración y la invasión de células ESCC; miR-301 elevado o PTEN reducido promueve la migración y la invasión de células ESCC. un Las capacidades de migración e invasión de las células KYSE-30 transfectadas evaluadas usando el ensayo Transwell después de la regulación a la baja de miR-310 o la regulación al alza de PTEN; b resultados estadísticos de las células KYSE-30 migradas e invasivas a través del ensayo Transwell después de la regulación a la baja de miR-310 o la regulación al alza de PTEN; c las capacidades de migración e invasión de las células Eca109 entre los grupos evaluados usando el ensayo Transwell después de la regulación al alza de miR-310 o la regulación a la baja de PTEN; d resultados estadísticos de las células Eca109 migradas e invasivas a través de la regulación positiva de miR-310 del ensayo Transwell o la regulación negativa de PTEN. * P <0,05 frente al grupo inhibidor-NC; & P <0,05 frente al grupo oe-NC; # P <0,05 frente al grupo inhibidor de miR-301 + si-NC; a P <0,05 frente al grupo imitador-NC; b P <0,05 frente al grupo si-NC; c P <0.05 versus el grupo miR-301 mimic + oe-NC, N =3. Los datos se expresaron como media ± desviación estándar y se utilizó ANOVA para las comparaciones entre múltiples grupos
El miR-301 inhibido o el PTEN sobreexpresado inducen la detención del ciclo celular y la apoptosis de las células ESCC; MiR-301 elevado o PTEN reducido suprime la detención del ciclo celular y la apoptosis de las células ESCC
Se utilizó citometría de flujo para detectar la transición del ciclo celular y la apoptosis de las células después de la transfección, y los resultados indicaron que en la línea celular KYSE-30 (Fig. 5a-d), la transfección del inhibidor de miR-301 o oe-PTEN promovió la tasa de apoptosis y aumento de células en la fase G0 / G1, mientras que disminuyó en las fases S y G2 / M; si-PTEN podría revertir la alteración de la apoptosis y la detención del ciclo celular inducida por el inhibidor de miR-301.
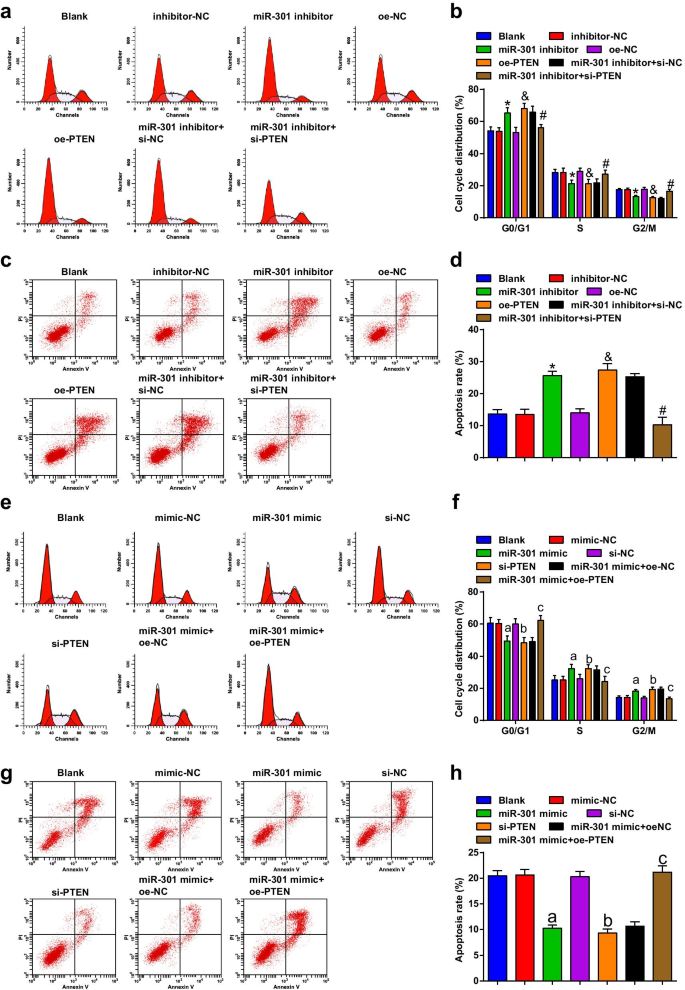
El miR-301 inhibido o el PTEN sobreexpresado inducen la detención del ciclo celular y la apoptosis de las células ESCC; miR-301 elevado o PTEN reducido suprime la detención del ciclo celular y la apoptosis de las células ESCC. un La distribución del ciclo celular de las células KYSE-30 en cada grupo se detectó mediante citometría de flujo después de la regulación a la baja de miR-310 o la regulación al alza de PTEN; b resultados estadísticos del porcentaje en las fases G0 / G1, S y G2 / GM de las células KYSE-30 en citometría de flujo después de la regulación a la baja de miR-310 o la regulación al alza de PTEN; c la apoptosis de las células KYSE-30 se detectó mediante citometría de flujo después de la regulación a la baja de miR-310 o la regulación al alza de PTEN; d tasa de apoptótica de células KYSE-30 transfectadas detectadas usando citometría de flujo después de regulación negativa de miR-310 o regulación positiva de PTEN; e la distribución del ciclo celular de las células Eca109 en cada grupo se detectó mediante citometría de flujo regulación positiva de miR-310 o regulación negativa de PTEN; f resultados estadísticos del porcentaje en las fases G0 / G1, S y G2 / GM de las células Eca109 en la citometría de flujo después de la regulación al alza de miR-310 o la regulación a la baja de PTEN; g la apoptosis de las células Eca109 se detectó mediante citometría de flujo después de la regulación por aumento de miR-310 o regulación por disminución de PTEN; h tasa de apoptótica de células Eca109 transfectadas detectadas usando citometría de flujo después de la regulación al alza de miR-310 o la regulación a la baja de PTEN. * P <0,05 frente al grupo inhibidor-NC; & P <0,05 frente al grupo oe-NC; # P <0,05 frente al grupo inhibidor de miR-301 + si-NC; a P <0,05 frente al grupo imitador-NC; b P <0,05 frente al grupo si-NC; c P <0.05 versus el grupo miR-301 mimic + oe-NC, N =3. Los datos se expresaron como media ± desviación estándar y se utilizó ANOVA para las comparaciones entre múltiples grupos
De acuerdo con los resultados de la citometría de flujo, hemos encontrado que en la línea celular Eca109 (Fig. 5e-h), la transfección de miR-301 imita o si-PTEN inhibe la tasa de apoptosis, disminuye las células en la fase G0 / G1 y aumenta la de la Fase S y fases G2 / M; La sobreexpresión de PTEN revirtió el efecto del imitador de miR-301 sobre la tasa de apoptosis y la detención del ciclo celular de las células Eca109. A partir de estos resultados, llegamos a la conclusión de que miR-301 regulado negativamente o PTEN regulado positivamente promovió la transición del ciclo celular y la apoptosis en las células ESCC, mientras que miR-301 inhibido o PTEN silenciado ejercía efectos opuestos.
El miR-301 inhibido o PTEN sobreexpresado restringe el crecimiento tumoral y la angiogénesis in vivo en ESCC; MiR-301 elevado o PTEN reducido aumenta el crecimiento tumoral y la angiogénesis in vivo en ESCC
En cada grupo se observó el crecimiento y los cambios de los tumores ESCC en ratones desnudos. Se evaluó el crecimiento tumoral y los resultados implicaron que en la línea celular KYSE-30 (Fig. 6a-e), los ratones desnudos inyectados con inhibidor de miR-301 o oe-PTEN redujeron el volumen y el peso del tumor; el papel represivo del inhibidor de miR-301 en el crecimiento tumoral fue abolido por si-PTEN. En la línea celular Eca109 (Fig. 6f-j), el volumen y el peso del tumor avanzaron en ratones desnudos inyectados con miR-301 imitador o si-PTEN; la sobreexpresión de PTEN revirtió el efecto del imitador de miR-301 sobre el crecimiento tumoral. Mientras tanto, la expresión de CD34 en xenoinjertos de ratones desnudos se evaluó mediante tinción inmunohistoquímica y los hallazgos mostraron que (Fig. 7a-d) en xenoinjertos KYSE-30, MVD se restringió después de regulación negativa de miR-301 o regulación positiva de PTEN; PTEN silenciado revirtió el impacto de la inhibición de miR-301 en MVD. En los xenoinjertos de Eca109, la MVD aumentó después de la regulación positiva de miR-301 o la regulación negativa de PTEN; el aumento de MVD inducido por miR-301 regulado positivamente podría ser abolido por PTEN sobreexpresado. Estos datos indicaron que la inhibición de miR-301 o la sobreexpresión de PTEN reprimieron el crecimiento tumoral y la angiogénesis en ESCC, mientras que la elevación de miR-301 o el silenciamiento de PTEN tuvieron efectos inversos.

Inhibited miR-301 or overexpressed PTEN restrains tumor growth in ESCC; elevated miR-301 or reduced PTEN increases tumor growth in ESCC. un Representative figures for the tumor growth observed by subcutaneous tumorigenesis in nude mice after KYSE-30 cells were transfected; b–d changes of tumor volume of each group after KYSE-30 cells were transfected; e changes of tumor weight of each group after KYSE-30 cells were transfected; f representative figures for the tumor growth observed by subcutaneous tumorigenesis in nude mice after Eca109 cells were transfected; g–i changes of tumor volume of each group after Eca109 cells were transfected; j changes of tumor weight of each group after Eca109 cells were transfected. * P < 0.05 versus the inhibitor-NC group; & P < 0.05 versus the oe-NC group; # P < 0.05 versus the miR-301 inhibitors + si-NC group; a P < 0.05 versus the mimic-NC group; b P < 0.05 versus the si-NC group; c P < 0.05 versus the miR-301 mimic + oe-NC group, n = 3 mice. Data were expressed as mean ± standard deviation, and ANOVA was used for comparisons among multiple groups

Inhibited miR-301 or overexpressed PTEN restrains angiogenesis in ESCC; elevated miR-301 or reduced PTEN increases angiogenesis in ESCC. un Representative images of tumor tissues observed by immunohistochemical staining in nude mice after KYSE-30 cells were transfected; b comparisons of MVD of KYSE-30 in tumor tissues among the groups; c representative images of tumor tissues observed by immunohistochemical staining in nude mice after Eca109 cells were transfected; d comparisons of MVD of Eca109 in tumor tissues among the groups *P < 0.05 versus the inhibitor-NC group; & P < 0.05 versus the oe-NC group; # P < 0.05 versus the miR-301 inhibitors + si-NC group; a P < 0.05 versus the mimic-NC group; b P < 0.05 versus the si-NC group; c P < 0.05 versus the miR-301 mimic + oe-NC group, n = 3 mice. Data were expressed as mean ± standard deviation, and ANOVA was used for comparisons among multiple groups
Discusión
EC is a kind of invasive malignancy in the gastrointestinal tract [16]. As the major type of EC, ESCC is a malignant tumor occurring in esophageal epithelial cells [17]. The miRNAs, known as small non-coding RNAs, have been demonstrated to function as a significant roles in leading molecules in the silencing of RNA [18]. Our research was designed to explore the effects of miR-301 and its target gene PTEN on ESCC progression, and we have found that the inhibited miR-301 could suppress angiogenesis and cell growth in ESCC by elevating PTEN.
MiR-301 expression was assessed, and we found that miR-301 was highly expressed in ESCC cells in comparison with HEEC, and the higher expression of miR-301 has also been found in ESCC tissues in contrast to the adjacent normal tissues. Similar to this result, Li et al. have identified that miR-301 presented high expression in myocardial infarction tissues [19]. In addition, we have elucidated that PTEN was targeted by miR-301, and the target relation has been pointed out by an extant literature [20]. We have also discovered that PTEN, which has been affirmed to be targeted by miR-301, was downregulated in both ESCC tissues and cells. Similarly, a previous research has unearthed that PTEN was poorly expressed in ESCC compared with non-tumor esophageal epithelial tissue [21]. Furthermore, Ma et al. have illuminated that PTEN expression was degraded in Eca109 cell line [22], which has also been selected for a series of experiments in this research. These studies provide evidence for the high expression of miR-301 and low expression of PTEN in ESCC.
Another important outcome in this research indicated that the inhibited miR-301 could repress the colony formation ability as well as the cell proliferation of ESCC cells via enhancing the PTEN expression, and elevated miR-301 or reduced PTEN had contrary effects. Similarly, Han et al. have elucidated that the downregulation of miR-301 mediated by luteolin has the ability to restrain the cell proliferation in prostate cancer [6]. A recent literature has revealed that the overexpression of PTEN suppresses the proliferation of pancreatic cancer cells [23], and a same result has been summarized in a study focusing on prostate cancer [24]. Besides, we have also unearthed that the downregulation of miR-301 or the elevation of PTEN could inhibit migration and invasion of ESCC cells, and elevated miR-301 or reduced PTEN exhibited the opposite trends. In accordance with this outcome, Shi et al. have supported that inhibited miR-301 attenuated migration and invasion of breast cancer cells [10], and it has been reported that the migration and invasion of ESCC cells could be repressed by the inhibition of miR-130b and the elevation of PTEN [25]. These publications helped verifying the oncogenic role of miR-301 and tumor-repressive effect of PTEN in diverse human cancers. Another result in our research was that inhibited miR-301 overexpressed PTEN to promote cell apoptosis and induce cell cycle arrest at the G0/G1 phase in ESCC cells, and elevated miR-301 or reduced PTEN had the inverse results. Similarly, it has been uncovered by a recent literature that activated PTEN induces cell cycle arrest and apoptosis in ESCC [26]. Consistently, Tian et al. have found in their study that the elevation of PTEN inhibited the angiogenesis by reducing the expression of vascular endothelial growth factor in hepatocellular carcinoma [27]. Based on the above data, the roles of miR-301 and PTEN in cell apoptosis and angiogenesis in diverse diseases were further confirmed. Consequently, we concluded that the downregulation of miR-301 could restrain the tumor growth in ESCC through the high expression of PTEN, and the similar conclusion has also been unveiled in breast cancer [10] and prostate cancer [28]. On the contrary, miR-301 elevation or PTEN reduction induced the tumor growth in ESCC. It could be concluded that miR-301 and PTEN participated in the in vivo cancer cell growth.
Conclusion
In this study, we have shown that the repression of miR-301 prohibits angiogenesis, cell proliferation, migration and invasion but promotes apoptosis in ESCC cells by upregulating PTEN. This research may further the understanding on potential molecular mechanisms of ESCC and provide novel targets for ESCC treatment.
Abreviaturas
- ESCC:
-
Carcinoma de células escamosas de esófago
- PTEN:
-
Phosphatase and tensin homologue
- MVD:
-
Densidad de microvasos
- EC:
-
Esophageal cancer
- miRNAs:
-
MicroRNAs
- LNM:
-
Lymph node metastasis
- UICC:
-
Union for International Cancer Control
- RT-qPCR:
-
Reacción en cadena de la polimerasa cuantitativa con transcripción inversa
- 3′UTR:
-
3′-Untranslated region
- WT:
-
Wild type
- MUT:
-
Mutant type
- HRP:
-
Peroxidasa de rábano picante
- FBS:
-
Suero fetal bovino
- OE:
-
Overexpressed
- NC:
-
Control negativo
- CCK-8:
-
Cell counting kit
- DMEM:
-
Dulbecco’s modified Eagle medium
- PI:
-
Yoduro de propidio
- FITC:
-
Fluorescein isothiocyanate
- MVD:
-
Densidad de microvasos
- ANOVA:
-
Analysis of variance
Nanomateriales
- La regulación a la baja del microARN-342-5p o la regulación al alza de Wnt3a inhibe la angiogénesis y mantiene la estabilidad de la placa aterosclerótica en ratones con aterosclerosis
- Preparación de nanopartículas de mPEG-ICA cargadas con ICA y su aplicación en el tratamiento del daño celular H9c2 inducido por LPS
- Análisis de actina y organización de la adhesión focal en células U2OS en nanoestructuras poliméricas
- Las nanofibras jerárquicas de poliamida 6-ZnO antibacteriana fabricadas por deposición de capas atómicas y crecimiento hidrotermal
- Estudio in vitro de la influencia de nanopartículas de Au en las líneas celulares HT29 y SPEV
- Promoción del crecimiento celular SH-SY5Y mediante nanopartículas de oro modificadas con 6-mercaptopurina y un péptido penetrante de neuronas
- Las nanopartículas de maghemita actúan como nanozimas, mejorando el crecimiento y la tolerancia al estrés abiótico en Brassica napus
- Nanopartículas de oro de forma redonda:efecto del tamaño de partícula y la concentración sobre el crecimiento de la raíz de Arabidopsis thaliana
- Soporte de catalizador anódico novedoso para pila de combustible de metanol directo:caracterizaciones y rendimiento de una sola pila
- Crecimiento propio de MOCVD y fotoluminiscencia mejorada espectacularmente de nanocables de núcleo-carcasa de InGaAs / InP
- 5G y el desafío del crecimiento exponencial de datos