


El microARN-133a exosómico derivado de células madre mesenquimatosas de la médula ósea restringe la fibrosis miocárdica y la transición epitelio-mesenquimatosa en ratas con miocarditis viral mediante la supresión de MAML1
Resumen
La miocarditis es una enfermedad caracterizada por una inflamación localizada o difusa del miocardio sin un tratamiento eficaz. Este estudio exploró el mecanismo regulador del microARN-133 (miR-133) secretado por el exosoma derivado de células madre mesenquimales de la médula ósea (BMSC-Exo) en la fibrosis miocárdica y la transición epitelio-mesenquimatosa (EMT) en ratas con miocarditis viral (VMC) mediante la regulación Mente maestra 1 (MAML1). Se aislaron y cultivaron BMSC en ratas para identificar su fenotipo inmunológico y capacidad osteogénica y adipogénica, y se extrajeron e identificaron BMSC-Exo. Los exosomas se obtuvieron mediante ultracentrifugación, los cuales se identificaron mediante microscopio electrónico de transmisión y análisis de transferencia Western. A las ratas se les inyectó el virus Coxsackie B3 para la preparación del modelo VMC, y los cardiomiocitos se aislaron, cultivaron y agruparon de la misma manera que los experimentos con animales (NC Exo , Ad-miR-133a Exo , Adas-miR-133a Exo ). Se llevaron a cabo experimentos in vivo e in vitro para determinar las funciones de miR-133a exosomal y MAML1 en la inflamación, apoptosis, EMT, fibrosis y viabilidad celular. La relación de dirección entre miR-133a y MAML1 se verificó mediante el ensayo del gen indicador de luciferasa dual. BMSC-Exo aumentó la expresión de miR-133a en ratas VMC y mejoró eficazmente la función cardíaca de la rata VMC y la fibrosis miocárdica, aumentó la viabilidad de los cardiomiocitos e inhibió el proceso de EMT. El miR-133a elevado en los exosomas fortaleció las mejoras. El miR-133a silenciado revirtió efectivamente los efectos de BMSC-Exo en ratas VMC. miR-133a apuntó a MAML1. La inhibición de MAML1 mejoró la función cardíaca y la fibrosis miocárdica en ratas VMC y podría revertir el efecto de los exosomas silenciados con miR-133a en ratas VMC. Nuestro estudio sugiere que el miR-133a exosómico elevado suprime la fibrosis miocárdica y la EMT en ratas con VMC mediante la regulación a la baja de MAML1, lo que inhibe la progresión de la miocarditis.
Introducción
La miocarditis se considera una enfermedad inflamatoria de las células del músculo cardíaco [1]. La miocarditis es obviamente más familiar en el hombre que en la mujer [2]. La miocarditis viral (CVM) es un factor principal que conduce a la miocardiopatía dilatada (MCD) y la muerte súbita en los jóvenes [3]. El rendimiento clínico de la miocarditis es variado, desde estados asintomáticos con signos y síntomas ambiguos hasta una destrucción miocárdica grave por virus y células inmunes que padecen shock cardiogénico y arritmias [1]. La miocarditis puede ser inducida por una variedad de elementos infecciosos, que consisten en virus, bacterias, clamidia, rickettsia, hongos y protozoos, junto con inductores no infecciosos. Entre las cuales, la infección viral ha sido la causa más común, especialmente en niños [4]. El virus Coxsackie B3 (CVB3), como el virus más importante que conduce a la miocarditis, puede provocar una respuesta al estrés oxidativo y apoptosis en la patogenia de la VMC, pero aún no se ha informado de un tratamiento especial de la VMC [5]. Además, la patogenia de la VMC no está bien documentada y también falta un tratamiento clínico exacto [3]. Por lo tanto, se necesitan con urgencia nuevos objetivos para mejorar el pronóstico de la enfermedad.
Los microARN (miARN) son ARN endógenos no codificantes que pueden regular la expresión de genes que codifican proteínas [6]. MiR-133a, como uno de los miARN específicos para el corazón, está implicado en el desarrollo del corazón y en algunas enfermedades cardiovasculares, que contienen infarto de miocardio (IM) [7]. Además, miR-133a es el que se expresa de forma aberrante en la miocardiopatía crónica por enfermedad de Chagas [8]. Además, el nivel de miR-133a en el miocardio está relacionado con la inflamación, la función del ventrículo izquierdo y el resultado clínico de la miocardiopatía inflamatoria [9]. Se encontraron miARN en exosomas derivados de mastocitos humanos y de ratón [10]. Los exosomas, vesículas de tamaño nanométrico liberadas por la mayoría de los tipos de células, se encuentran en diferentes fluidos biológicos [11]. Los exosomas pueden transferir su carga a las células receptoras, lo que se ha demostrado que cambia la composición bioquímica y las vías de señalización de las células receptoras [12, 13]. La evidencia ha demostrado que los miARN exosomales modificados están relacionados con la patogenia de la miocarditis inducida por CVB3 [14]. Se ha abordado que el miR-125b-5p exosómico de células madre mesenquimales de la médula ósea condicionadas por hipoxia reduce la apoptosis de los cardiomiocitos y mejora la reparación cardíaca isquémica [15]. Además, el miR-25-3p exosómico de las CMM alivia el IM al reducir la apoptosis de los cardiomiocitos y la respuesta inflamatoria [16]. Es de interés que el miR-133a expresado en forma cardíaca exosómica esté conectado a la troponina-I de la troponina cardíaca [17]. Mastermind-like 1 (MAML1) fue un gen descendente de miR-133a sometido a cribado cruzado en nuestro estudio que, según se ha informado, implica una lesión por isquemia / reperfusión miocárdica (I / R) [18]. Además, un estudio reciente ha mencionado que la eliminación de MAML1 posee la función antifibrótica en la fibrosis hepática [19].
Iluminado por estudios previos, se pregunta si el miR-133a exosómico derivado de BMSC podría mediar en la miocarditis. Por lo tanto, este estudio se inició con la hipótesis de que el miR-133 transportado por el exosoma derivado de BMSC (BMSC-Exo) mejora la fibrosis miocárdica y la transición epitelial-mesenquimatosa (EMT) en ratas VMC mediante la regulación de MAML1.
Materiales y métodos
Aprobación ética
El estudio fue autorizado por el Comité Institucional de Uso y Cuidado Animal del Cuarto Hospital Afiliado de la Facultad de Medicina de la Universidad de Zhejiang. Los animales fueron tratados con humanidad.
Aislamiento de BMSC
Los animales experimentales fueron ratas macho Sprague-Dawley (SD) maduras libres de patógenos específicos (SPF) de grado (Centro de Animales Experimentales de la Facultad de Medicina de la Universidad de Zhejiang, Zhejiang, China). Las ratas se sacrificaron mediante inyección intraperitoneal con pentobarbital sódico y se esterilizaron con alcohol al 75%. Se extrajeron el fémur y la tibia en una mesa ultralimpia, se extrajeron el tejido muscular y conjuntivo y la cavidad de la médula se enjuagó repetidamente con medio de Eagle modificado de Dulbecco bajo en glucosa (DMEM). El líquido se centrifugó para recoger los precipitados que se resuspendieron y se incubaron durante 24 h (el medio se cambió cada 2-3 días). Al crecer hasta la fase logarítmica, las BMSC se separaron con tripsina al 0,25% (Gibco, Carlsbad, California, EE. UU.), Se centrifugaron y se resuspendieron en una solución de cultivo de MSC (Cyagen Biosciences Inc., Guangzhou, China). La suspensión se pasó a una proporción de 1:2. Se repitió la operación anterior y se utilizaron las BMSC del cuarto pase para experimentos posteriores.
Identificación de BMSC
Los antígenos de superficie de las BMSC de cuarto paso en crecimiento logarítmico se identificaron mediante citometría de flujo. Las BMSC se separaron con tripsina al 0,25% (1 ml) que contenía ácido etilendiaminotetraacético, se centrifugaron, se resuspendieron con solución salina tamponada con fosfato (PBS) apropiada y se centrifugaron a 151 g. A continuación, las BMSC se resuspendieron con PBS que contenía suero bovino fetal (FBS) reciente al 2% (Gibco) para preparar una suspensión de una sola célula. Se incubaron los anticuerpos monoclonales FITC-CD34, PE-CD29 y PE-CD44 (5 μL cada uno, BD Biosciences, Franklin Lakes, NJ, EE. UU.) Con la suspensión celular (100 μL), se centrifugaron a 151 g y se resuspendieron con 500 μL de PBS que contiene paraformaldehído al 1% y se fija durante 30 min. Los marcadores de fondo se identificaron mediante el uso de anticuerpos monoclonales del control de homotipo.
Citometría de flujo:la suspensión unicelular se fijó y centrifugó a 151 g. Luego, las BMSC se resuspendieron con PBS que contenía paraformaldehído al 1%, se probaron en el citómetro de flujo MACS Quart y se analizaron con el software correspondiente.
Inducción de osteogénesis y adipogénesis de BMSC
Las BMSC en el cuarto pase se sembraron en placas de 6 pocillos con 200 células / ml. La solución de inducción de osteoblastos y la solución de inducción adipogénica (Cyagen Biosciences Inc.) se agregaron en BMSC de 60 a 70% de confluencia. Las BMSC en otros dos pozos no se agregaron con líquidos de inducción como controles. Las BMSC se indujeron durante 14 días y se fijaron con paraformaldehído al 4%. Luego, los osteoblastos y adipocitos diferenciados se implementaron con tinción con rojo de alizarina y tinción con rojo de aceite O (Wuhan Pulande Biological Technology Co., Ltd., Wuhan, China), y se observaron bajo un microscopio.
Aislamiento e identificación de exosomas
Las BMSC en el cuarto paso se cultivaron durante 48 h para recolectar el sobrenadante que luego se centrifugó (800 gy 2000 g), se filtró con membranas de filtro de 0,22 μm y 100.000 MW y se centrifugó (100.000 g) para recoger los precipitados. Luego, los precipitados se resuspendieron con PBS, se centrifugaron nuevamente a 100,00 g para obtener la precipitación del exosoma. La suspensión de BMSC-Exo en PBS se sometió a detección de concentración mediante ácido bicinconínico (BCA) y detección de proteína productora de exosomas (CD63, CD81 y CD9) mediante análisis de transferencia Western (Proteintech, Chicago, IL, EE. UU.).
La infección por adenovirus recombinante media la modificación del gen miR-133a de las BMSC
Las BMSC se pasaron durante la noche. El control normal (cantidad igual de PBS), el control negativo de miR-133a (NC), la sobreexpresión de miR-133a (Ad-miR-133a) y la baja expresión de miR-133a (Adas-miR-133a) ( Hanbio Biotechnology Co., Ltd., Shanghai, China) se transfectaron con BMSC en línea con 100 multiplicidad de infección (MOI). Se cultivaron las BMSC y los exosomas correspondientes (NC Exo , NC Exo , Ad-miR-133a Exo y Adas-miR-133a Exo ) se obtuvieron mediante ultracentrifugación [20].
Establecimiento del modelo VMC en ratas y agrupación de animales experimentales
Se dividieron ratas SD macho maduras de grado SPF en 10 grupos, con ocho ratas cada uno. El Coxsackievirus B3 (CVB3) fue proporcionado por el Instituto de Biotecnología Médica de la Academia China de Ciencias Médicas (Beijing, China).
Se inyectó por vía intraperitoneal CVB3 (10 mg / kg) en ratas mientras se inyectaba PBS o BMSC-Exo (100 µg) a través de la vena de la cola. Se inyectaron ratas normales para los controles con solución de cultivo CVB3 y PBS. Las ratas inyectadas con 10 mg / kg de CVB3 se inyectaron adicionalmente con PBS, MSC exo , NC Exo , Adas-miR-133a Exo , Ad-miR-133a Exo , si-NC o si-MAML1 (RIBOBIO, Guangzhou, China).
Las ratas se inyectaron continuamente durante 7 días y se obtuvo la sangre del globo ocular. La sangre se centrifugó y el suero se recogió, se subenvasó y se almacenó a -20 ° C. Después de la eutanasia de las ratas, se tomaron muestras de corazón, se fijaron con formaldehído al 10%, se deshidrataron con alcohol en gradiente, se aclararon con xileno y se embebieron con parafina, seccionadas para observación histológica. Una parte de las secciones se colocaron a -80 ° C como materiales de experimentos de biología molecular.
Ecocardiografía
El séptimo día después de la inyección del virus, se inyectó a las ratas por vía intraperitoneal 25 mg / kg de pentobarbital sódico. Después de la anestesia completa, el cable de la extremidad de la máquina de electrocardiograma conectado con la aguja del electrodo se insertó por vía subcutánea en los extremos de las extremidades de las ratas y se registró el electrocardiograma del cable de la extremidad. Luego, las ratas se fijaron en decúbito supino ligeramente hacia la izquierda, se depiló el tórax y se conectó el electrocardiograma de derivación II para obtener el espectro Doppler del pulso del flujo sanguíneo de la aorta en la sección cardíaca paraesternal de cuatro cámaras. Los indicadores incluyeron el grosor de la pared posterior del ventrículo izquierdo (LVPW), el diámetro telesistólico del ventrículo izquierdo (LVID), la fracción de acortamiento del ventrículo izquierdo (FS) y la fracción de eyección del ventrículo izquierdo (FEVI).
Tinción con hematoxilina-eosina (HE)
Los tejidos se fijaron con paraformaldehído al 4%, se deshidrataron, aclararon y embebieron con parafina. Luego se desparafinaron las secciones de 4 μm, se tiñeron con hematoxilina (Servicebio, Wuhan, China), se diferenciaron con alcohol ácido clorhídrico al 1%, se volvieron a azul y se tiñeron con eosina, se deshidrataron, se aclararon con xileno, se sellaron con goma neutra, y observado bajo un microscopio óptico (Olympus, Tokio, Japón).
Tinción de colágeno de Masson
Las secciones de parafina se desparafinaron, se tiñeron con hematoxilina durante menos de 2 min, se tiñeron con una solución de Lichun magenta y se enjuagaron rápidamente con una solución de ácido acético glacial al 0,5%. Luego, las secciones se tiñeron con una solución acuosa de fosfato de aluminio al 1%, se tiñeron de rojo oscuro a rojo brillante a rosa y se observaron al microscopio. Luego, las secciones se tiñeron con azul de anilina (pulanda), se deshidrataron convencionalmente con xileno y se sellaron. Se utilizó el software de análisis de imágenes médicas Image-Proplus 6.0 para medir el área de tinción positiva de las fibras de colágeno y la fracción de volumen de colágeno (CVF) =área de colágeno / área de campo total. Se distinguió la ubicación de la tinción y el color de las fibras de colágeno (los cardiomiocitos eran rojos y las fibras de colágeno tenían bandas azules o estructuras homogéneas en el espacio intercelular).
Tinción de marcación de extremos de desoxinucleotidil transferasa mediada por desoxiuridina trifosfato-biotina Nick (TUNEL)
Las secciones de parafina se desparafinaron, se colocaron en el tampón de citrato y se hornearon a 350 W durante 10 min. Las secciones se agregaron con 50 μL de solución de TUNEL, se unieron con 50 μL de agente de conversión-peroxidasa, se desarrollaron con DAB y se observaron al microscopio. Las secciones se colocaron en hematoxilina, se sumergieron en etanol I-II al 95%, se unieron con etanol anhidro I-II, xileno I-II y se sellaron. Los resultados se analizaron con un microscopio óptico.
Ensayo inmunoabsorbente ligado a enzimas (ELISA)
El factor de necrosis tumoral α (TNF-α), la interleucina (IL) -1β e IL-6 se detectaron mediante kits de ELISA (BOSTER Biological Technology Co. Ltd., Wuhan, China). La sangre del globo ocular se centrifugó a 604 g para recoger el suero superior. El sobrenadante obtenido por centrifugación del medio de cultivo celular se detectó en el experimento celular. Había siete gradientes de concentración en el estándar de dilución de muestras. El pocillo del blanco se unió con diluyente de muestra y se añadió otro pocillo con tetrametilbencidina (TMB), se colocaron dos pocillos duplicados para cada concentración. Los pocillos de muestras se unieron con 50 μL de diluyente de muestra y las muestras a su vez. Cada pocillo se hizo reaccionar con 100 μL de anticuerpo primario (excepto el pozo TMB) durante 1 h, así como con 300 μL de solución salina tamponada con tris (TBS) 0.01 M y 100 μL de solución de trabajo del complejo de avidina-biotina-peroxidasa (excepto el pozo TMB ). Luego, a cada pocillo se le añadieron 300 μL de TBS 0,01 M y se incubó con 100 μL de TMB. El valor de densidad óptica (DO) y la concentración de cada pocillo se midieron inmediatamente y se trazó la curva estándar.
Reacción en cadena de la polimerasa cuantitativa con transcripción inversa (RT-qPCR)
La expresión de MiR-133a, colágeno Ι, colágeno III, α-SMA, TGF-β1, CTGF, E-cadherina y FSP-1 en tejidos miocárdicos y cardiomiocitos se detectó mediante RT-qPCR. El ARN total se extrajo de cardiomiocitos o tejidos miocárdicos y se transcribió inversamente a ADNc mediante un kit de extracción de ARN (Takara, Dalian, China), y los cebadores de RT-PCR se sintetizaron a través de Invitrogen (Guangzhou, China); las secuencias se muestran en la Tabla 1. La expresión génica cuantitativa relativa se analizó utilizando gliceraldehído-3-fosfato deshidrogenasa (GAPDH) o U6 como genes de control de carga de acuerdo con 2 - △△ Ct método.
Análisis de Western Blot
Las ratas se sacrificaron con anestesia. Los tejidos del miocardio se congelaron y se trituraron en nitrógeno líquido. A continuación, se mezcló la solución madre de fenilmetanosulfonilfluoruro de inhibidores de proteasa con tampón de lisis celular en una proporción de 1:100 (Beyotime Biotechnology Co., Ltd., Shanghai, China). Las muestras se lisaron con una solución mixta y se extrajeron las proteínas de las células. La concentración de proteína total se detectó con el kit BCA. Las muestras se mezclaron con tampón de carga 5X a 4:1, se implementaron con baño de agua hirviendo durante 10 min, se bañaron con hielo y se centrifugaron. Se realizó la separación por electroforesis y las proteínas se transfirieron a una membrana de fluoruro de polivinilideno (Servicebio) con una solución de transferencia eléctrica. Luego, la membrana se bloqueó con leche desnatada en polvo al 5% y se unió con los anticuerpos primarios CD63, CD81 y CD9 (anticuerpos policlonales de conejo anti-rata de Proteintech, 1:100), MAML1 (ab65090, Abcam, MA, EE. UU., 1:1000) y GAPDH (Santa Cruz Biotechnology, Inc, Santa Cruz, CA, EE. UU., 1:1000). Luego, la membrana se goteó con el anticuerpo secundario, IgG marcada con peroxidasa de rábano picante (Cell Signaling Technology, Beverly, MA, EE. UU., 1:1000) y se sumergió en una solución de reacción de quimioluminiscencia mejorada (Pierce, Rockford, IL, EE. UU.). Con GAPDH como control de carga, las imágenes de impresión de proteínas se analizaron con el software ImageJ2x.
Cultivo y pasaje de cardiomiocitos
Se tomaron ratas SD de 3-5 días (Centro de Animales Experimentales de la Facultad de Medicina de la Universidad de Zhejiang, Zhejiang, China). La parte ventricular se enjuagó con solución salina equilibrada de Hank preenfriada, se cortó en trozos pequeños y se separó con tripsina al 0,25%. Las piezas se añadieron con una cantidad adecuada de medio completo al 10% para la terminación del desprendimiento y se centrifugaron a 151 g. Se aplicó DMEM que contenía FBS al 20% para resuspensión celular. Los cardiomiocitos se purificaron mediante un método de adherencia diferencial, y la tasa de supervivencia se observó mediante tinción con azul tripán, los cardiomiocitos supervivientes se cultivaron. A las 24 h, los cardiomiocitos se adhirieron a la pared y empezaron a pulsar. Después de 72 h, los pseudópodos se expandieron.
Construcción del modelo VMC de cardiomiocito
Los cardiomiocitos en el cuarto pase en la fase de crecimiento logarítmico fueron seleccionados e infectados con MSC Exo , NC Exo , Adas-miR-133a Exo y Ad-miR-133a Exo . La solución de virus 100 Tcid50 CVB3 (100 µl) se añadió a las células para inducir un modelo de VMC celular. Al mismo tiempo, se agregó la misma cantidad de solución de mantenimiento a las células para un control, y los exosomas correspondientes se unieron en cardiomiocitos después de 1 h de infección para un cultivo de 47 h.
Ensayo del kit de recuento celular (CCK) -8
Se aplicó el kit de detección de células CCK-8 (Beyotime) para detectar la tasa de supervivencia de los cardiomiocitos. Cuando crecieron a la fase logarítmica, las células se separaron con tripsina al 0,25% y se sembraron en una placa de cultivo celular de 96 pocillos a 2,5 × 10 4 células / por pocillo. Junto con la solución CCK-8 (10 μL / pocillo), las células se cultivaron continuamente durante 1 a 4 h, y la DO 450 nm El valor se midió mediante un lector de microplacas.
Citometría de flujo
Se aplicó el método de doble tinción de AnnexinV-APC / yoduro de propidio (PI) para detectar la apoptosis celular. Las células se centrifugaron, se resuspendieron con 250 μl de tampón de unión (4 ml de tampón de unión + 12 ml de agua desionizada) y se ajustaron a 1 x 10 6 células / mL. La suspensión celular de 100 μL se agregó con 5 μL de Anexina V-APC (BD Biosciences) y 5 μL de solución de PI (BD Biosciences), se cargó en el citómetro de flujo y se analizó automáticamente en una computadora.
Ensayo génico indicador de luciferasa dual
La secuencia de tipo salvaje (wt) o de tipo mutante (mut) de la región no traducida 3 (UTR) de MAML1 se clonó en el vector pGL3-M (Promega, WI, EE. UU.), Luego MAML1-3-UTR-wt o MAML1- Se generaron 3-UTR-mut. Los vectores, junto con miR-133a mimic o NC, se cotransfectaron en cardiomiocitos a través de Lipofectamine 2000. La actividad de luciferasa se probó 48 h después mediante el sistema de genes indicadores de luciferasa dual (Promega) [21]
Ensayo de inmunoprecipitación de ARN (RIP)
Se utilizó el kit RIP (Millipore, EE. UU.) Para detectar la unión de MAML1 y miR-133a. Las células se lisaron mediante tampón de ensayo de radioinmunoprecipitación (P0013B, Beyotime, Shanghai, China), se centrifugaron a 1400 gy se incubaron con anticuerpos para coprecipitar. Se resuspendieron perlas magnéticas (50 μL) en 100 μL de tampón de lavado RIP y se incubaron con 5 g de anticuerpo anti-MAML1 (1 g / mL, ab155786) o IgG (1:100, ab172730). El complejo de perlas magnéticas-anticuerpo se resuspendió en 900 μL de tampón de lavado RIP, se interactuó con 100 μL de extracto celular, se digirió con proteinasa K y se detectó mediante RT-qPCR [22].
Análisis estadístico
Se aplicó el software estadístico SPSS 21.0 (IBM Corp. Armonk, NY, EE. UU.) Para el análisis de los datos. Los datos de medición se expresaron como media ± desviación estándar. La prueba t se aplicó a las comparaciones entre dos grupos. Se utilizó el análisis de varianza unidireccional (ANOVA) para la comparación entre grupos y la prueba post hoc de Tukey para las comparaciones por pares. Los predictores se mantuvieron si eran significativos en un P valor de 0.05 o menor.
Resultados
Identificación de BMSC y BMSC-Exo
Microscópicamente, las BMSC eran fusiformes y redondeadas, y estaban adheridas a la pared en un patrón de vórtice o radial (Fig. 1A). Después de la tinción con rojo aceite O de la inducción de la adipogénesis, las gotitas de lípidos de la cuarta BMSC eran rojas y había gotitas de lípidos redondas de diferentes tamaños (Fig. 1B). Después de la inducción de la osteogénesis, las células que expresan los nódulos calcificados mostraron enrojecimiento después de la tinción con rojo de alizarina y una distribución desigual de los nódulos calcificados y las células superpuestas (Fig. 1C). La citometría de flujo manifestó que se expresaban los marcadores de MSC CD29 y CD44 (> 95%), pero no se expresaba el antígeno de superficie de células madre hematopoyéticas CD34 (<95%) (Fig. 1D). Estos resultados manifestaron que las BMSC eran de alta pureza y se ajustaban a los estándares de MSC de la Sociedad Internacional de Terapia Celular.
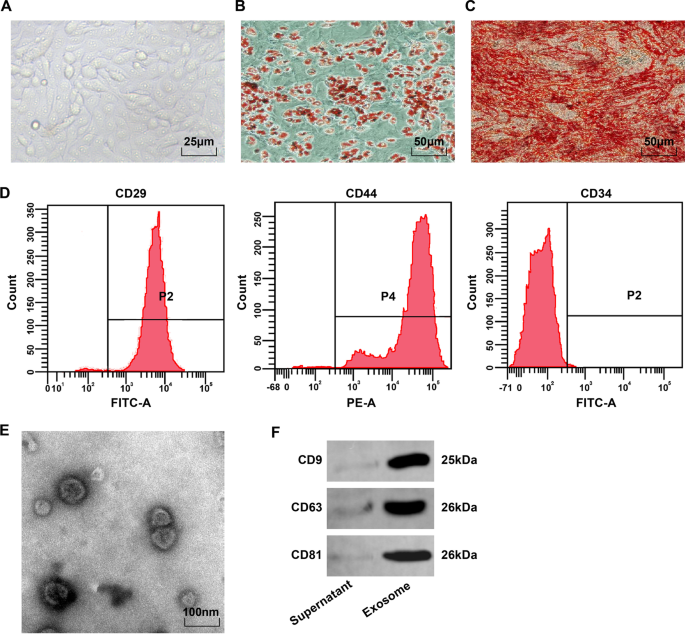
Observación del fenotipo de BMSC e identificación de BMSC-Exo. A Observación morfológica de BMSC en el cuarto pasaje; B Los resultados de la tinción con rojo aceite O de los adipocitos; C Los resultados de la tinción con rojo de alizarina de los osteoblastos; D El fenotipo de BMSC detectado mediante citometría de flujo. E Observación de microscopio electrónico de BMSC-Exo; F Bandas de proteínas de CD9, CD63 y CD81
El microscopio electrónico de transmisión observó que BMSC-Exos eran vesículas ovaladas con estructura membranosa periférica clara, diferentes tamaños y un diámetro de 40-100 nm (Fig. 1E). El análisis de transferencia Western demostró que los productos extraídos que expresaban el exosoma originaban proteínas características de CD9, CD63 y CD81 (Fig. 1F).
El miR-133a exosómico elevado mejora los síntomas de la miocarditis
Se observó la transfección de BMSC con adenovirus recombinante miR-133a (Fig. 2A). Se observó un gran número de expresión de fluorescencia verde de NC, Ad-miR-133a y Adas-miR-133a bajo el microscopio de fluorescencia invertido, lo que indica que el vector de adenovirus recombinante podría transfectar eficazmente BMSC. Para probar la eficacia de transfección de miR-133a, se midió la expresión de miR-133a en BMSC y sus exosomas mediante RT-qPCR. Se descubrió que la regulación positiva de miR-133a aumentaba la expresión de miR-133a mientras que la regulación negativa de miR-133a disminuía la expresión de miR-133a (Fig. 2B). Posteriormente, inyectamos exosomas que contenían miR-133a en ratas. Bajo el microscopio de fluorescencia invertido, se observó expresión de fluorescencia verde en ratas VMC después del tratamiento de NC Exo , Ad-miR-133a Exo , o Adas-miR-133a Exo , lo que indica que el vector de adenovirus recombinante infectó tejidos miocárdicos de ratas (Fig. 2C). El experimento RT-qPCR también encontró que la expresión de miR-133a en ratas VMC aparentemente estaba disminuida; La expresión de miR-133a fue obviamente elevada en ratas VMC inyectadas con Ad-miR-133a Exo pero disminuyó en ratas VMC inyectadas con Adas-miR-133a Exo (Figura 2D). En cuanto a las condiciones generales de las ratas, se observó que las condiciones generales de las ratas control normales eran normales, y las características de VMC aparentemente se expresaron en ratas VMC y ratas VMC inyectadas con Adas-miR-133a Exo , como cabello áspero y desordenado, disnea y poca dieta. En ratas VMC tratadas con MSC Exo , NC Exo y Ad-miR-133a Exo , estos signos se mejoraron en diferentes grados. El peso de las ratas VMC se redujo continuamente desde 1 día después de la infección y la inyección de MSC Exo , NC Exo , o Ad-miR-133a Exo aumentó el peso de las ratas. El peso de las ratas VMC tratadas con Ad-miR-133a Exo se incrementó obviamente y el peso de las ratas VMC inyectadas con Adas-miR-133a Exo se redujo claramente (Fig. 2E).
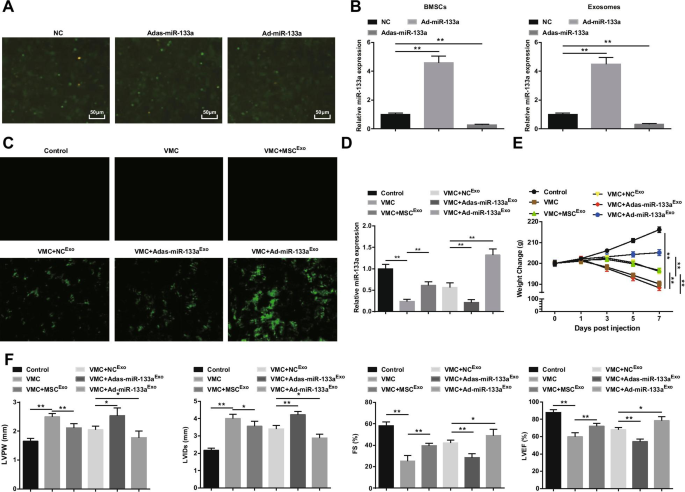
El miR-133a exosómico regulado al alza alivia la miocarditis. A Transfección con BMSC de adenovirus recombinante miR-133a; B Detección por RT-qPCR de la expresión de miR-133a en BMSC y sus exosomas después de regular miR-133a; C eficacia de transfección de miR-133a probada mediante microscopio de fluorescencia invertido; D La expresión relativa de miR-133a en tejidos miocárdicos analizados mediante RT-qPCR; E Cambio de peso de las ratas en cada grupo; F Determinación de LVPW, LVIDs, FS y LVEF en ratas de cada grupo. * P <0,05; ** P <0,001
La observación de la función miocárdica sugirió que (Fig. 2F), las ratas VMC tenían un aumento de LVPW y LVID, y una disminución de FS y LVEF. Después de la inyección de exosomas, disminuyó LVPW y LVID, y obviamente se mostró FS y LVEF elevadas en ratas VMC. Adas-miR-133a Exo tratamiento deteriorado mientras Ad-miR-133a Exo función miocárdica mejorada en ratas VMC.
El miR-133a exosómico regulado positivamente inhibe la inflamación en los tejidos miocárdicos de ratas VMC
La tinción con HE manifestó que las fibras del miocardio en las ratas de control normales estaban dispuestas de manera cercana y no había infiltración de células inflamatorias en el mesénquima. Los cardiomiocitos en las ratas VMC estaban desorganizados y el mesénquima estaba infiltrado por un gran número de células inflamatorias. Los cardiomiocitos en ratas VMC inyectadas con MSC Exo o NC Exo estaban ordenadas, con una pequeña cantidad de células inflamatorias infiltrándose en el mesénquima. Los cardiomiocitos en ratas VMC después de Adas-miR-133a Exo el tratamiento se organizó de forma desordenada y se infiltraron las células inflamatorias del mesénquima. Los cardiomiocitos en ratas VMC tratadas con Ad-miR-133a Exo se dispusieron ordenadamente sin infiltración evidente de células inflamatorias (Fig. 3A).
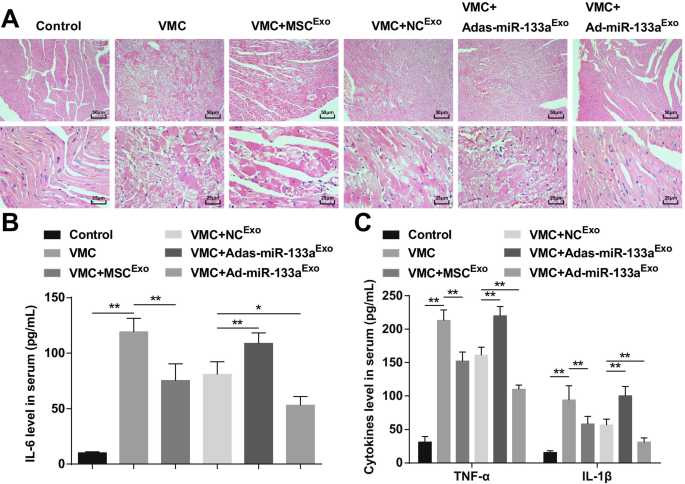
El aumento de miR-133a exosómico frena la inflamación en los tejidos del miocardio con VMC. A Tinción HE de tejido miocárdico de rata en cada grupo; B La expresión de IL-6 en suero analizada mediante ELISA; C La expresión de TNF-α e IL-1β en suero analizada mediante ELISA. * P <0,05; ** P <0,001
ELISA indicó que (Fig. 3B, C) los factores inflamatorios (TNF-α, IL-1β e IL-6) estaban obviamente aumentados en ratas VMC. Las ratas VMC inyectadas con Ad-miR-133a Exo había reducido los niveles de factores inflamatorios. Adas-miR-133a Exo el tratamiento provocó factores inflamatorios elevados en ratas VMC.
El miR-133a exosómico elevado disminuye la CVF en los tejidos miocárdicos de ratas con VMC
La tinción de Masson reveló que las fibras miocárdicas en ratas normales estaban dispuestas de manera cercana, casi sin fibras colágenas azules. Después de la inyección de CVB3, los cardiomiocitos estaban hipertróficos, con hiperplasia del tejido conectivo y un gran número de fibras colágenas azules, y la CVF estaba obviamente elevada. Tratados con exosomas, los cardiomiocitos se ordenaron, disminuyó la hiperplasia del tejido conjuntivo intercelular, las fibras de colágeno azul y la CVF disminuyeron claramente. El espacio intercelular del miocardio de ratas VMC inyectadas con Adas-miR-133a Exo se ensanchó, las células obviamente aumentaron de tamaño, las fibras de colágeno azul y el CVF aumentaron claramente; el espacio intercelular se redujo, la distribución de fibras de colágeno azul y el CVF se redujeron en ratas VMC con Ad-miR-133a Exo tratamiento (Fig. 4A, B).
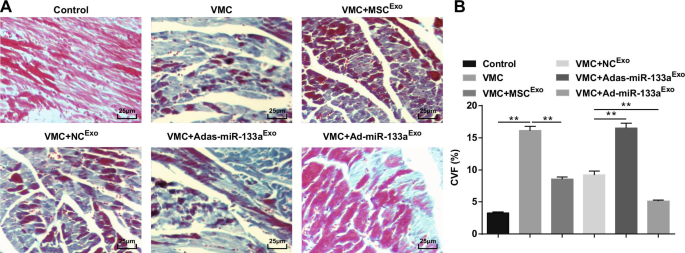
El miR-133a exosómico regulado al alza disminuye la CVF en los tejidos miocárdicos de ratas con VMC. A Tinción de Masson de tejidos miocárdicos en ratas; B Fracción de volumen de colágeno de ratas en cada grupo. ** P <0,001
El aumento de miR-133a exosómico reduce la expresión de colágeno I, colágeno III, TGF-β1 y CTGF en tejidos miocárdicos de ratas con VMC
El colágeno I y el colágeno III son los componentes principales del colágeno, que se distribuyen principalmente en las uniones celulares y las membranas celulares, la sustancia intercelular y el citoplasma. TGF-β1 y CTGF son las proteínas características de la fibrosis. Los hallazgos de RT-qPCR demostraron que los niveles de expresión de ARNm de colágeno I, colágeno III, TGF-β1 y CTGF aumentaron en ratas VMC, pero disminuyeron después del tratamiento con exosomas. Las ratas VMC tratadas con Ad-miR-133a Exo tenían niveles de expresión de ARNm disminuidos de colágeno I, colágeno III, TGF-β1 y CTGF, mientras que las ratas VMC después de Adas-miR-133a Exo el tratamiento mostró la situación opuesta (Fig. 5A-C).
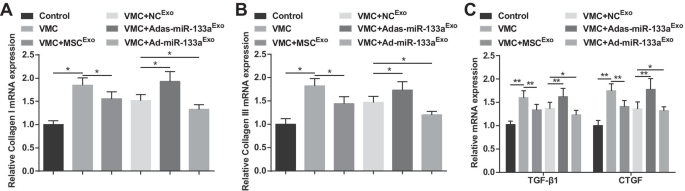
El miR-133a exosómico elevado reduce la expresión de ARNm de colágeno I, colágeno III, TGF-β1 y CTGF en tejidos miocárdicos de ratas con VMC. A La expresión de ARNm de colágeno I en tejidos miocárdicos de ratas se detectó mediante RT-qPCR; B La expresión de ARNm de colágeno III en tejidos de miocardio de ratas se detectó mediante RT-qPCR; C La expresión de ARNm de TGF-β1 y CTGF en tejidos miocárdicos de ratas se detectó mediante RT-qPCR. * P <0,05; ** P <0,001
El miR-133a exosómico regulado positivamente inhibe la apoptosis de cardiomiocitos en tejidos miocárdicos de ratas con VMC
La tinción TUNEL mostró que los cardiomiocitos apoptóticos eran de color negro pardusco o amarillo pardusco con condensación nuclear. El número de células apoptóticas aumentó en ratas VMC, lo que se atenuaría mediante el tratamiento con exosomas. Las ratas VMC inyectadas con Ad-miR-133a Exo había reducido el número de células apoptóticas y las inyectadas con Adas-miR-133a Exo había aumentado el número de células apoptóticas (Fig. 6A, B).
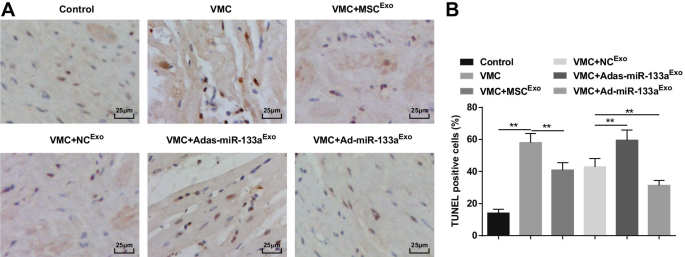
El aumento de miR-133a exosómico inhibe la apoptosis de cardiomiocitos en tejidos miocárdicos de ratas con VMC. A Tinción TUNEL de tejidos miocárdicos de rata en cada grupo; B El número de células TUNEL positivas en cada grupo. ** P <0,001
El miR-133a exosómico elevado deprime la EMT en los tejidos miocárdicos de ratas con VMC
E-cadherina, α-SMA y FSP-1 son indicadores clave de EMT. Los resultados de RT-qPCR demostraron que los niveles de expresión de ARNm de α-SMA y FSP-1 estaban elevados y el nivel de expresión de ARNm de E-cadherina disminuyó en ratas VMC. Además, los niveles de expresión de ARNm de α-SMA y FSP-1 se redujeron y el nivel de expresión de ARNm de E-cadherina aumentó en ratas VMC después del tratamiento con exosomas. Los niveles de expresión de ARNm de α-SMA y FSP-1 se elevaron y el nivel de expresión de ARNm de E-cadherina disminuyó en ratas VMC tratadas con Adas-miR-133a Exo , mientras que la expresión de estos indicadores fue opuesta en ratas VMC inyectadas con Ad-miR-133a Exo (Fig. 7A, B).
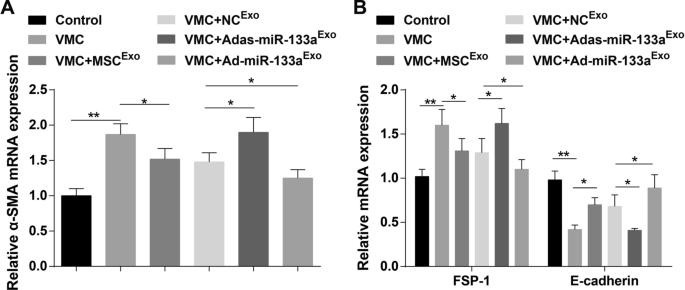
El miR-133a exosómico regulado positivamente reprime la EMT en los tejidos miocárdicos de ratas con VMC. A La expresión de ARNm de α-SMA en tejidos de miocardio de rata en cada grupo se detectó mediante RT-qPCR; B La expresión de ARNm de FSP-1 y E-cadherina en tejidos de miocardio de rata en cada grupo se detectó mediante RT-qPCR. * P <0,05; ** P <0,001
El miR-133a exosómico regulado al alza deprime la inflamación de los cardiomiocitos en VMC
Como resultado, la microscopía de fluorescencia capturó la expresión fluorescente verde en ratas VMC tratadas con NC Exo , Ad-miR-133a Exo , o Adas-miR-133a Exo , lo que indica que el vector de adenovirus recombinante infectó cardiomiocitos de ratas (Fig. 8A). RT-qPCR y ELISA descubrieron que (Fig.8B, D) la expresión de miR-133a se redujo y los factores inflamatorios (TNF-α, IL-1β e IL-6) aumentaron en ratas VMC, lo que se revertiría con el tratamiento con exosomas . Las ratas VMC tratadas con Ad-miR-133a Exo tenían miR-133a regulado al alza y factores inflamatorios reducidos en ratas VMC, mientras que las tratadas con Adas-miR-133a Exo presentó una disminución de miR-133a y un aumento de los niveles de factores inflamatorios en ratas VMC.
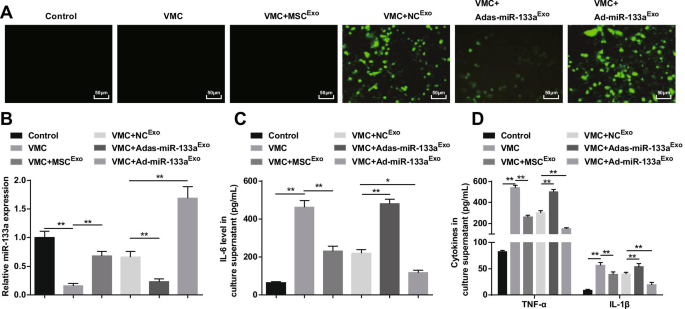
El miR-133a exosómico elevado frena la inflamación de los cardiomiocitos en VMC. A eficacia de transfección de miR-133a probada mediante microscopio de fluorescencia invertido; B La expresión relativa de miR-133a en cardiomiocitos de ratas en cada grupo; C Expresión de IL-6 en el sobrenadante de cultivo de cardiomiocitos en cada grupo; D Expresión de TNF-α e IL-1β en el sobrenadante de cultivo de cardiomiocitos en cada grupo. * P <0,05; ** P <0,001
El miR-133a exosómico elevado promueve la viabilidad celular y reprime la apoptosis de cardiomiocitos en VMC
La apoptosis y la viabilidad celular se detectaron mediante doble tinción con AnnexinV-APC / PI y ensayo CCK-8. Los resultados revelaron que hubo un aumento obvio en la tasa de apoptosis, una disminución en la viabilidad celular de los cardiomiocitos en ratas VMC. El tratamiento con exosomas redujo la tasa de apoptosis y mejoró la viabilidad de los cardiomiocitos. Adas-miR-133a Exo mejoró la tasa de apoptosis e interrumpió la viabilidad de los cardiomiocitos en ratas VMC. Ad-miR-133a Exo El tratamiento funcionó con efectos opuestos en los cardiomiocitos de ratas VMC (Fig. 9A-C).
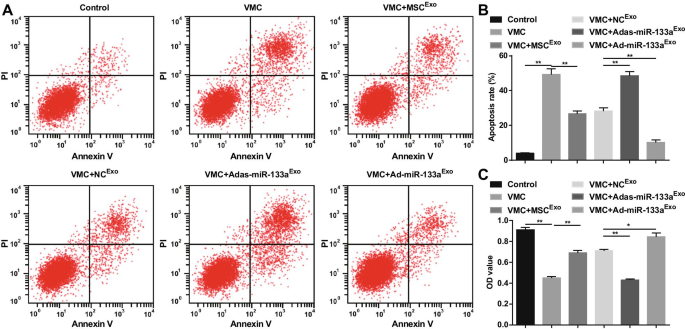
El aumento de miR-133a exosómico promueve la viabilidad y reprime la apoptosis en cardiomiocitos en VMC. A La apoptosis de los cardiomiocitos se detecta mediante citometría de flujo; B Resultados de cuantificación de A; C La viabilidad celular detectada mediante el ensayo CCK-8. * P <0,05; ** P <0,001
miR-133a apunta a MAML1
Se ha informado de que el miARN-193b regulado al alza reduce el daño I / R del miocardio al dirigirse a MAML1 [18]. Basándonos en eso, hicimos un cribado cruzado de genes descendentes de miR-133a a través de los sitios web de bioinformática PITA, miRanda, PicTar, microT y miRmap, y seleccionamos MAML1 como objetivo de miR-133a (Fig. 10A). Construimos MAML1-wt o MAML1-mut, y cardiomiocitos cotransfectados con miR-133a mimic o NC. Los resultados mostraron que el mimético de miR-133a redujo la actividad luciferasa de MAML1-wt (Fig. 10B). El experimento RIP verificó además la relación de dirección entre miR-133a y MAML1 (Fig. 10C). La detección de RT-qPCR y Western blot de la expresión de MAML1 mostró que la expresión de MAML1 estaba disminuida en los cardiomiocitos transfectados con el mimético de miR-133a (Fig. 10D, E).
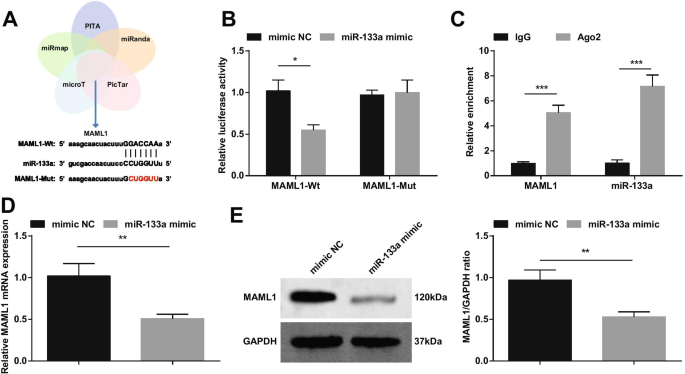
miR-133a apunta a MAML1. A los objetivos de miR-133a predichos en los sitios web de bioinformática; B La relación de dirección entre miR-133a y MAML1 verificada por el experimento del gen indicador de luciferasa dual; C La relación de dirección entre miR-133a y MAML1 verificada por el experimento RIP; D / E La expresión de MAML1 cambia después de la regulación positiva de miR-133a detectada por RT-qPCR y Western blot. * P <0,05; ** P <0,01; *** P <0,001
La inhibición de MAML1 tiene un efecto protector en ratas con miocarditis y revierte el efecto de exosomas inhibidos por miR-133a en ratas con VMC
Para estudiar más a fondo el efecto de MAML1 regulado por miR-133a en ratas con VMC, inyectamos adenovirus si-MAML1 o si-NC en ratas VMC o ratas VMC que habían sido tratadas con exosomas silenciados con miR-133a. El éxito de la inyección fue validado por RT-qPCR y Western blot (Fig. 11A, B). Los resultados manifestaron que la inyección de si-MAML1, el peso de las ratas VMC aumentó (Fig. 11C), la función cardíaca mejoró (Fig. 11D-G), la patología del tejido miocárdico y la fibrosis se atenuaron (Fig. 12A-C), Se inhibieron la inflamación sérica (Fig. 12D, E) y la apoptosis de los cardiomiocitos (Fig. 13A-G). Además, los efectos nocivos de los exosomas silenciados con miR-133a en ratas VMC se revirtieron después de la inyección de si-MAML1.
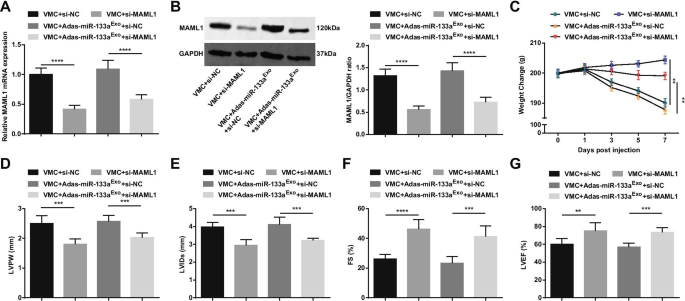
La inhibición de MAML1 tiene un efecto protector en ratas con miocarditis y puede revertir el efecto de exosomas silenciados con miR-133a en ratas con VMC. A / B Expresión de MAML1 en tejido miocárdico de ratas detectada por RT-qPCR y Western blot; C. Cambio de peso de las ratas; D - G Determinación de LVPW, LVID, FS y LVEF en ratas; ** P <0,01; *** P <0,001; **** P <0,0001
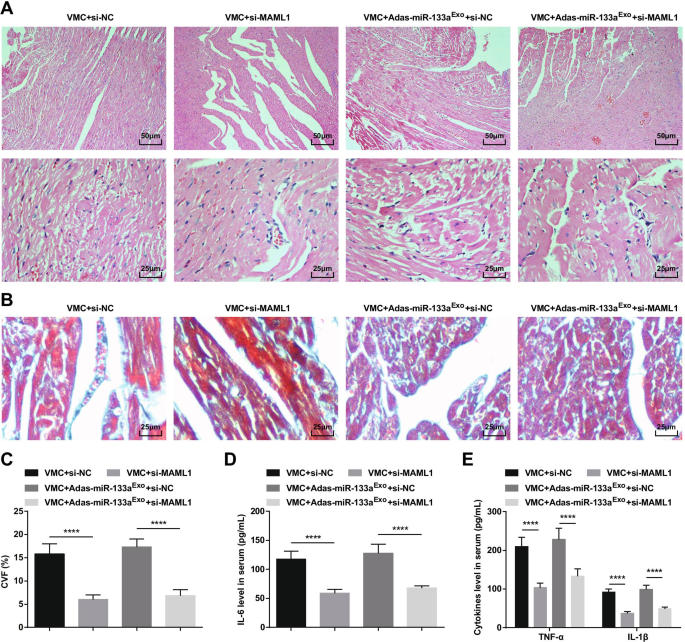
La inhibición de MAML1 puede revertir el efecto de los exosomas silenciados con miR-133a en ratas con VMC. A Tinción HE de tejido miocárdico de rata; B Tinción de Masson de tejidos miocárdicos en ratas; C CVF de ratas; D La expresión de IL-6 en suero analizada mediante ELISA; E La expresión de TNF-α e IL-1β en suero analizado mediante ELISA. **** P <0,0001
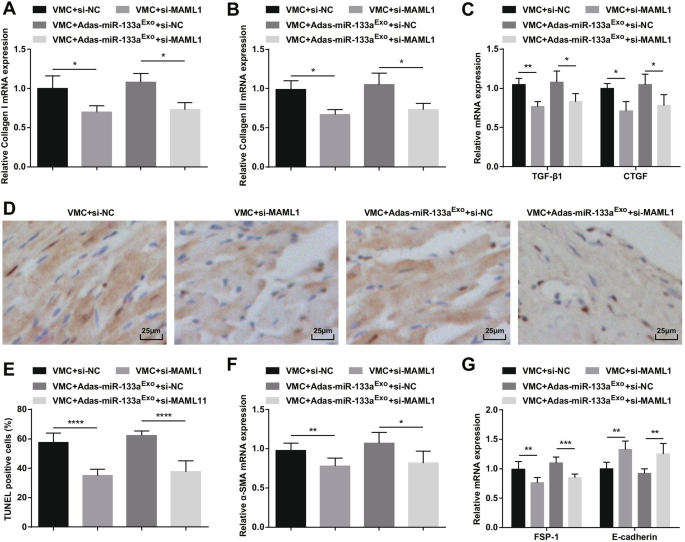
La inhibición de MAML1 puede revertir el efecto de los exosomas inhibidores de miR-133a en ratas con VMC. A La expresión de ARNm de colágeno I en tejidos miocárdicos de ratas se detectó mediante RT-qPCR; B La expresión de ARNm de colágeno III en tejidos de miocardio de ratas se detectó mediante RT-qPCR; C La expresión de ARNm de TGF-β1 y CTGF en tejidos miocárdicos de ratas se detectó mediante RT-qPCR; D Tinción TUNEL de tejidos miocárdicos de rata en cada grupo; E El número de células TUNEL positivas en cada grupo. F La expresión de ARNm de α-SMA en tejidos de miocardio de rata en cada grupo se detectó mediante RT-qPCR; G La expresión de ARNm de FSP-1 y E-cadherina en tejidos de miocardio de rata en cada grupo se detectó mediante RT-qPCR. * P <0,05; ** P <0,01; **** P <0,0001
Discusión
La miocarditis es una enfermedad cardíaca inflamatoria que produce MCD e insuficiencia cardíaca y es inducida con mayor frecuencia por infecciones virales como el CVB3 [2]. Un estudio ha revelado que miR-133 alivia la apoptosis de los cardiomiocitos y la remodelación eléctrica en ratones con VMC [23]. Además, los miARN exosomales modificados también se relacionan con la patogenia de la miocarditis inducida por CVB3 [14]. Los exosomas derivados de células progenitoras cardíacas facilitan la apoptosis inducida por CVB3 mediante la restricción de la proliferación de CVB3 en VMC [24]. Este estudio exploró el mecanismo regulador del miR-133 exosómico derivado de BMSC en la fibrosis miocárdica y la EMT en ratas VMC (archivo adicional 1:Fig. 1).
El estudio encontró que la expresión de miR-133a se redujo en VMC. Como se demostró antes, la expresión de miR-133a está disminuida en MI [7]. Un estudio también ha sugerido que la expresión relativa de miR-133 en corazones de ratón de la VMC obviamente disminuye con el contraste con los controles [23]. Hay algunas conexiones de miARN con exosomas. La expresión diferencial de los exosomas y de los miARN exosomales en la enfermedad se ha considerado como biomarcadores de la enfermedad con la realización de un diagnóstico clínico no invasivo junto con sus potenciales terapéuticos [25]. Lin y otros . han descubierto que miR-133 se clasifica especialmente en exosomas derivados de células progenitoras endoteliales humanas causadas por hipoxia / reoxigenación (H / R) para aumentar la angiogénesis de fibroblastos y la EMT [26]. Otro estudio ha revelado que las CMM exhiben una comunicación con las células del parénquima cerebral y pueden modular el crecimiento de neuritas mediante la transferencia de miR-133b a las células neurales a través de exosomas [27].
El principal hallazgo de este trabajo manifestó que miR-133a exosómico regulado positivamente promovió la viabilidad celular, inhibió la inflamación, apoptosis, EMT y fibrosis en ratas con VMC. Se adaptan bien a una investigación anterior de que el silencio del miR-133a revierte los polisacáridos del astrágalo inhibición de la proliferación celular del osteosarcoma MG63 inducida por el tratamiento, junto con la promoción de la apoptosis celular [28]. Otro estudio ha revelado que el miR-133a sobreexpresado suprime la angiogénesis, la apoptosis, la fibrosis y la inflamación, mientras que acelera la remodelación cardíaca terapéutica en las enfermedades isquémicas del miocardio [29]. Similar a nuestro estudio, Li et al . han declarado que miR-133 inhibe la apoptosis de los cardiomiocitos regulando la expresión de genes relacionados con la apoptosis en los corazones de ratones VMC [23]. Se ha informado que el miR-133a sobreexpresado deprime la apoptosis inducida por hipoxia y fortalece la supervivencia de los cardiomiocitos [30]. Mientras tanto, los exosomales miR-30a y miR-181d séricos regulados al alza pueden tener el potencial de ser aplicados como biomarcadores para el diagnóstico de VMC [14].
Otro hallazgo en nuestro estudio fue que miR-133a exosómico regulado positivamente disminuyó la CVF, redujo la expresión de colágeno I y colágeno III en ratas con VMC. Un artículo ha aclarado que la liberación del factor 18 de crecimiento de fibroblastos de una membrana de colágeno provoca la actividad osteoblástica que participa en miR-133a regulado a la baja [31]. La expresión excesiva in vitro de miR-133a deprime la hipertrofia de los cardiomiocitos y reduce la expresión de colágeno [32], como se evidencia en otro estudio. La CVF es igual a la relación entre el área de colágeno y la suma del área del miocardio y el área de colágeno, y el valor medio muestra la CVF de la sección [33]. Este hallazgo también fue informado por Wang et al . que el modelo de ratones VMC se construye con éxito por la infección por CVB3, que manifiesta una expresión aparentemente más alta de CVF en contraste con el grupo de control [34]. Además, el hallazgo es consistente con el de Ferreira et al . quien demuestra que miR-133a puede asumir un papel importante en la modulación de la expresión génica en la patogénesis de la miocardiopatía crónica de la enfermedad de Chagas, con un vínculo potencial como herramientas de diagnóstico y pronóstico [8]. Además, la evidencia ha demostrado que la eliminación de MAML1 puede reducir la hipertrofia de los cardiomiocitos pretratados [35]. En nuestro estudio, encontramos que MAML1 era el gen diana de miR-133a y la inhibición de MAML1 revirtió los efectos de los exosomas silenciados con miR-133a en ratas con VMC. En la lesión por isquemia-reperfusión miocárdica, la regulación a la baja de MAML1 mediada por miR-193b podría reducir en parte el infarto y las enzimas del miocardio, así como atenuar la apoptosis de los cardiomiocitos [18]. Además, hay un informe que sugiere que la deficiencia de MAML1 podría aliviar la fibrogénesis hepática [19].
Conclusión
En conclusión, este estudio ofrece evidencia de que miR-133a está regulado a la baja en ratas con VMC, y el miR-133a exosómico elevado mejora la función cardíaca y restringe la fibrosis miocárdica y la EMT en ratas con VMC, así como también mejora la viabilidad y reprime la apoptosis de cardiomiocitos en VMC mediante la focalización en MAML1. Nuestro estudio también sugiere que la inhibición de MAML1 tiene un efecto protector en ratas con miocarditis e invierte el efecto de los exosomas inhibidos por miR-133a en ratas con VMC. La identificación del miR-133a exosómico en la fibrosis miocárdica y la EMT de la miocarditis puede ampliar potencialmente nuestra comprensión de los mecanismos que sustentan la miocarditis y también tener valor clínico como un nuevo objetivo molecular. Se deben realizar más investigaciones para avanzar en el tratamiento de esta enfermedad.
Abreviaturas
- miR-133:
-
MicroARN-133
- BMSC-Exo:
-
Exosoma derivado de células madre mesenquimales de la médula ósea
- EMT:
-
Transición epitelial-mesenquimal
- VMC:
-
Miocarditis viral
- CVF:
-
Fracción de volumen de colágeno
- DCM:
-
Miocardiopatía dilatada
- CVB3:
-
Virus Coxsackie B3
- miARN:
-
MicroARN
- MI:
-
Infarto de miocardio
- SPF:
-
Libre de patógenos específicos
- SD:
-
Sprague – Dawley
- DMEM:
-
Medio Eagle modificado de Dulbecco
- PBS:
-
Solución salina tamponada con fosfato
- FBS:
-
Suero fetal bovino
- NC:
-
Control negativo
- MOI:
-
Multiplicidad de infección
- LVPW:
-
Espesor de la pared posterior del ventrículo izquierdo
- LVID:
-
Diámetro telesistólico del ventrículo izquierdo
- LVEF:
-
Fracción de eyección del ventrículo izquierdo
- HE:
-
Hematoxilina-eosina
- TUNEL:
-
Etiqueta terminal de muesca de desoxiuridina trifosfato-biotina terminal mediada por desoxinucleotidil transferasa
- ELISA:
-
Ensayo inmunoabsorbente ligado a enzimas
- TMB:
-
Tetrametilbencidina
- TBS:
-
Solución salina tamponada con Tris
- ABC:
-
Complejo avidina-biotina-peroxidasa
- OD:
-
Densidad óptica
- RT-qPCR:
-
Reacción en cadena de la polimerasa cuantitativa con transcripción inversa
- GAPDH:
-
Gliceraldehído-3-fosfato deshidrogenasa
- CTGF:
-
Factor de crecimiento del tejido conectivo
- CCK:
-
Kit de recuento celular
- PI:
-
Yoduro de propidio
- ANOVA:
-
Análisis de varianza
Nanomateriales
- Nanopartículas de fósforo negro promueven la diferenciación osteogénica de EMSC a través de la expresión TG2 regulada hacia arriba
- Estructura y propiedades electrónicas de la nanoarcilla de caolinita dopada con metal de transición
- Fibras de carbón activado “densamente cubiertas” por Ag Nanohair mediante autoensamblaje y recocido térmico rápido
- Estudio de la fuerza de adhesión y la transición vítrea de películas delgadas de poliestireno mediante microscopía de fuerza atómica
- Estudios TEM y STEM sobre las morfologías transversales de películas antirreflectantes de SiO2 de banda ancha de doble / triple capa
- Un escrutinio comparativo in vivo de nanopartículas de óxido de zinc y cobre biosintetizadas por vías de administración intraperitoneal e intravenosa en ratas
- Desarrollo y caracterización de compuestos de vitrocerámica que contienen Sr basados en hidroxiapatita biogénica
- Materiales de la familia del grafeno en la regeneración del tejido óseo:perspectivas y desafíos
- Nanoscopía basada en microscopía de fuerza atómica de células madre derivadas de tejido adiposo humano que se diferencian condrogenicamente:nanoestructura e integrina expresión β1
- Células madre mesenquimales marcadas con nanopartículas de azul de Prusia:evaluación de la viabilidad celular, proliferación, migración, diferenciación, citoesqueleto y expresión de proteínas …
- Propiedades electrónicas de la butaca Nanocintas de fosforo negro modificadas en los bordes por los elementos de transición V, Cr y Mn