


El microARN-326-5p restaurado inhibe la apoptosis neuronal y atenúa el daño mitocondrial mediante la supresión de STAT3 en la isquemia cerebral / lesión por reperfusión
Resumen
Los estudios han explorado en gran medida el papel de los microARN (miARN) en la lesión por isquemia / reperfusión cerebral (IC / RI). Pero el mecanismo específico de miR-326-5p en CI / RI todavía es difícil de alcanzar. Por lo tanto, este estudio fue para desenmascarar el mecanismo del miR-326-5p / transductor de señal y activador del eje de transcripción-3 (STAT3) en CI / RI. Se establecieron dos modelos (privación de oxígeno y glucosa [OGD] en neuronas corticales primarias de rata y oclusión de la arteria cerebral media [MCAO] en ratas Sprague-Dawley) para imitar CI / RI in vitro e in vivo, respectivamente. Se realizaron ensayos de pérdida y ganancia de función con neuronas tratadas con OGD y con ratas MCAO. Posteriormente, se probaron la viabilidad, apoptosis, estrés oxidativo y potencial de membrana mitocondrial en neuronas tratadas con OGD, así como cambios patológicos, apoptosis y potencial de membrana mitocondrial en tejidos cerebrales de ratas MCAO. Se detectó la expresión de mitofusina-2 (Mfn2), miR-326-5p y STAT3 en neuronas tratadas con OGD y en tejidos cerebrales de ratas MCAO. Se redujeron Mfn2 y miR-326-5p, y STAT3 se elevó en neuronas tratadas con OGD y tejidos cerebrales de ratas MCAO. Expresión de STAT3 dirigida y regulada negativamente por miR-326-5p. Restaurar miR-326-5p o reducir STAT3 reforzó la viabilidad, inhibió la apoptosis y el estrés oxidativo, aumentó el potencial de membrana mitocondrial y aumentó la expresión de Mfn2 en neuronas tratadas con OGD. La regulación positiva de miR-326-5p o la regulación negativa de STAT3 alivió los cambios patológicos, inhibió la apoptosis y el potencial de membrana mitocondrial elevado y la expresión de Mfn2 en tejidos cerebrales de ratas con MCAO. Este estudio aclara que miR-326-5p regulado al alza o STAT3 regulado a la baja protege contra CI / RI al elevar la expresión de Mfn2.
Introducción
La lesión por isquemia / reperfusión cerebral (IC / RI) es un tipo de lesión cerebral seguida de un accidente cerebrovascular isquémico [1]. El rasgo más característico de la I / R cerebral es la isquemia cerebral transitoria inicial después de la reperfusión [2]. CI / RI activa la apoptosis neuronal y conduce al hipocampo y daño cortical [3]. En la actualidad, la aplicación clínica más común es trombolítica para IC / RI [4]. Sin embargo, la reperfusión aumentaría la producción de especies reactivas de oxígeno (ROS), lo que provocaría daño en el ADN intracelular, daño relacionado con el estrés oxidativo, oxidación de proteínas y peroxidación de lípidos, deteriorando aún más la barrera hematoencefálica y el edema [5]. Por lo tanto, la urgencia de buscar nuevas opciones específicas encabeza la prioridad de CI / RI.
Los microARN (miARN) se consideran ampliamente como roles potenciales y pronósticos en CI / RI [6, 7]. Por ejemplo, un artículo ha demostrado que miR-202-5p atenúa el daño neuronal y los déficits neurológicos, así como el daño celular inducido por privación de oxígeno y glucosa (OGD) en ratas modelo con oclusión de la arteria cerebral media (MCAO) en la lesión isquémica [8] . Además, se ha encontrado que la regulación positiva de miR-98 mejora los resultados neurológicos en ratones con accidente cerebrovascular I / R [9] y la sobreexpresión de miR-451 alivia la apoptosis cerebral isquémica en ratones con IC / RI [10]. Específicamente, miR-326-5p tiene capacidad proangiogénica para las células progenitoras endoteliales y podría mejorar la función cardíaca después de un infarto agudo de miocardio [11]. Sin embargo, el estudio que gira en torno a miR-326-5p en CI / RI aún está en su infancia. Los transductores de señal y el activador transcripcional (STAT) son una familia única de proteínas que es activada por CI / RI [12]. STAT3 se predijo como un objetivo de miR-326-5p en el sitio web de bioinformática; por lo tanto, estudiamos el papel de STAT3 mediado por miR-326-5p en CI / RI. STAT3 deteriora los procesos neuroinflamatorios isquémicos y la lesión cerebral secundaria al liberar mediadores proinflamatorios [13, 14]. Específicamente, la vía de la quinasa 2 activada por Janus (JAK2) / STAT3 desempeña un papel funcional en el bloqueo de la apoptosis celular inducida por CI / RI [15]. La mitofusina-2 (Mfn2) es un factor de fusión mitocondrial que podría proteger contra I / RI cardio-cerebrovascular [16] y mejorar la apoptosis neuronal inducida por hipoxia en el daño cerebral isquémico [17]. También se ha registrado que Mfn2 ejerce un efecto protector sobre CI / RI [18].
En una palabra, una investigación menos profunda ha descubierto el papel combinado de miR-326-5p y STAT3 en CI / RI. Dado eso, este estudio se lanza con la hipótesis de que miR-326-5p atenúa CI / RI a través de la orientación STAT3.
Materiales y métodos
Declaración de ética
Los experimentos fueron aprobados por el Comité de Uso y Cuidado de Animales del Hospital Afiliado de la Universidad de Yangzhou; Universidad de Yangzhou, y se llevó a cabo con la Guía para el Cuidado y Uso de Ratas Experimentales del Hospital Afiliado de la Universidad de Yangzhou; Universidad de Yangzhou.
Animales experimentales
Se mantuvieron ratas macho adultas Sprague-Dawley (de 6 a 8 semanas, 250 ± 30 g) del Centro de Medicina Comparada de la Universidad de Yangzhou (Yangzhou, China) en un entorno específico libre de patógenos con acceso libre a alimentos y agua. (24 ± 1) ° C, (50 ± 5)% de humedad y ciclo de luz / oscuridad de 12 h.
Identificación e aislamiento celular
Las ratas fueron sacrificadas por decapitación y sus cerebros se cortaron en 1 mm 3 para configurar suspensiones celulares, que se mezclaron con una solución de azul de tripán al 0,4% a 9:1 (la concentración final de azul de tripán fue de 0,04%) y se calculó la densidad celular y la tasa de supervivencia mediante un hemocitómetro. Las celdas (1 × 10 6 células / ml) se sembraron en un matraz de cultivo celular recubierto previamente con L-polilisina (0,1 mg / ml) a 5 × 10 6 células por matraz. Cuando las células se adhirieron a las paredes, se incubaron con el medio renovado (NeurobasalA + B27 + L-glutamina) y se cultivaron continuamente durante 6 a 7 días con el medio cambiado a la mitad cada 2 a 3 días.
Identificación celular:se utilizaron neuronas corticales de rata para preparar portaobjetos de neuronas que se incubaron con el anticuerpo policlonal primario de conejo anti-Nestina de rata (NES) (1:200), así como con la inmunoglobulina de cabra anti-conejo marcada con isotiocianato de fluoresceína (FITC). G (1:50, ambos de Cell Signaling Technology, Beverly, MA, EE. UU.). Posteriormente, las células se sellaron con un extintor anti-fluorescente y se observaron mediante un microscopio de fluorescencia.
Establecimiento del modelo OGD
Las neuronas se cultivaron en una solución de Earle sin glucosa y se expusieron a una mezcla de gas de 95% N 2 y 5% CO 2 . Después de 30 min de ventilación con presión positiva, se formó un ambiente hipóxico en el que se incubaron las células (simulación de isquemia por OGD in vitro). Después de 90 minutos de incubación, las células se incubaron con el medio celular de mantenimiento después de eliminar la solución de Earle sin glucosa. Las células se cultivaron de forma rutinaria para experimentos posteriores (simulación de reperfusión por reperfusión de oxígeno-glucosa in vitro). El medio normal bajo normoxia sirvió como control.
Las neuronas se dividieron en el grupo de control; el grupo OGD, el grupo de control negativo (NC) (neuronas tratadas con OGD transfectadas con oligonucleótidos mezclados), el grupo agomir miR-326-5p (neuronas tratadas con OGD transfectadas con agomir miR-326-5p), el grupo sh-STAT3 (Neuronas tratadas con OGD transfectadas con ARNhc de STAT3) y el grupo miR-326-5p agomir + sobreexpresión (oe) -STAT3 (neuronas tratadas con OGD transfectadas con miR-326-5p agomir y vector pcDNA-STAT3). Todos los oligonucleótidos y vectores fueron proporcionados por GenePharma (Shanghai, China).
Usando lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific), se transfectaron oligonucleótidos mezclados, miR-326-5p agomir, STAT3 shRNA o miR-326-5p agomir y pcDNA-STAT3 en neuronas primarias de rata y la eficacia de la transfección se probó mediante transcripción inversa cuantitativa reacción en cadena de la polimerasa (RT-qPCR) o Western blot después de 48 h.
Ensayo de bromuro de 3- (4, 5-dimetiltiazol-2-il) -2, 5-difeniltetrazolio (MTT)
Las neuronas se hicieron una suspensión a 1 × 10 6 células / mL y se cultivaron durante 0, 24 y 48 h, respectivamente. En un ambiente estéril, las neuronas se incubaron durante 2 a 4 h en una solución de MTT al 10% y se agregaron 150 μL de solución disuelta de Formanzan (dimetilsulfóxido) hasta que el cristal se disolvió lo suficiente. El valor de densidad óptica (DO) se midió en un lector de microplacas a 570 nm.
Ensayo de apoptosis por citometría de flujo
Las neuronas se separaron con tripsina al 0,25%, se centrifugaron a 2000 rpm y se resuspendieron en PBS. Luego, las neuronas se centrifugaron a 2000 rpm y el sedimento se resuspendió mediante tampón de unión y se incubó sucesivamente con 5 μL de anexina V-FITC y 5 μL de yoduro de propidio (PI). Se aplicó un citómetro de flujo para detectar la tasa de apoptosis.
Detección de potencial de membrana mitocondrial JC-1
Las células en la placa de 6 pocillos se enjuagaron con PBS, se les añadió 1 mL de medio de cultivo celular y 1 mL de solución de trabajo de tinción JC-1 (50 μL de JC-1, 8 mL de agua ultrapura y 2 mL de tampón de tinción JC-1) (SolarbioScience , Beijing, China) y se incubó durante 15 min. El tampón de tinción 1 × JC-1 se configuró con 4 ml de agua destilada y 1 ml de tampón de tinción JC-1. Después de la incubación, las células se tripsinizaron, se resuspendieron en tampón JC-1 y se probaron en un citómetro de flujo. Se calculó la proporción relativa de fluorescencia verde.
Detección de estrés oxidativo
Los niveles de ROS intracelulares se midieron con el kit de detección de diacetato de 2,7-diclorofluoresceína (DCF-DA) (Abcam). En resumen, las neuronas se separaron con tripsina al 0,25%, se resuspendieron y se incubaron con 10 μm de DCF-DA durante 30 min. Después de eso, se midió la fluorescencia de DCF con luz de excitación / emisión a 495 nm / 529 nm mediante espectroscopia de fluorescencia (BD Biosciences).
La detección de malondialchehyche intracelular (MDA), contenido de glutatión (GSH) y actividad de superóxido dismutasa (SOD) se detectó mediante colorimetría química utilizando kits de ensayo comerciales (Beyotime Biotechnology Co., Shanghai, China). El valor de DO se midió con un lector de microplacas.
Establecimiento modelo de rata
Las ratas se anestesiaron mediante inyección intraperitoneal de pentobarbital sódico (50 µg / kg). Se hizo una incisión en la línea media en el cuello, se ligó la carótida externa y se bloqueó la arteria carótida común con un clip arterial. Se insertó cuidadosamente una sutura de nailon monofilamento recubierta de silicona 3-0 en la arteria carótida interna hasta obtener una ligera resistencia. Después de 90 min de isquemia cerebral transitoria, se retiró la sutura y se realizó la reperfusión durante otras 24 h. El grupo de simulación recibió el mismo tratamiento, excepto que la sutura no entró en la arteria carótida interna. Durante la operación, se utilizó un lecho termostático para mantener la temperatura rectal a 37 ± 0,5 ° C [19].
Las ratas se dividieron en 6 grupos (n =6) y se les inyectó plásmido o miR en el cerebroventricular lateral antes del establecimiento de los modelos MCAO, incluido el grupo simulado, el grupo MCAO (ratas modeladas), el grupo NC (ratas modeladas con inyección cerebroventricular lateral de 5 μL de oligonucleótidos revueltos [100 μM]), el grupo de miR-326-5p agomir (ratas modeladas con inyección cerebroventricular lateral de 5 μL de miR-326-5p agomir [100 μM]), el grupo sh-STAT3 (ratas modeladas con inyección cerebroventricular lateral de 5 μL de vector de interferencia STAT3 [100 μM]) y el grupo miR-326-5p agomir + oe-STAT3 (ratas modeladas con inyección cerebroventricular lateral de 5 μL de miR-326-5p agomir [100 μM] y 5 μL Vector de sobreexpresión STAT3 [100 μM]) [20, 21].
Colección de muestras
Las ratas se sacrificaron mediante anestesia, se perfundieron los corazones y se obtuvieron los cerebros completos. Los tejidos con lesión cerebral se cortaron en 1 × 1 × 1 mm 3 masas de tejido para la preparación de cortes de parafina, RT-qPCR, análisis de transferencia Western y detección de potencial de membrana mitocondrial.
Detección de potencial de membrana mitocondrial en la corteza cerebral
Se separó la corteza cerebral de ratas SD y se cortó en 1 mm 3 . Los bloques de tejido se colocaron en una red de nailon de malla 300 y se les añadió PBS. Luego, la suspensión celular se centrifugó a 1000 r / min, se le añadió etanol al 70% y se centrifugó nuevamente a 1000 r / min. Posteriormente, la muestra se resuspendió en 1 mL de PBS y la suspensión (1 × 10 6 células, 100 μL) se hizo reaccionar con solución de colorante Rhodamine123 (10 μL, 5 μg / mL) y se analizó en un citómetro de flujo a 488 nm.
Tinción HE
El tejido de cerebro de rata se sumergió en paraformaldehído al 4% durante 24 h, y se prepararon secciones de parafina, se cortaron en 4 μm, se esparcieron en agua a 40 ° C y se hornearon a 60 ° C. Desparafinados e hidratados de forma rutinaria, las secciones de parafina se tiñeron con solución de tinción HE. Después de la tinción HE, las neuronas dañadas mostraron contracción del núcleo, edema celular, vacuolización y núcleos oscurecidos. Se utilizó el microscopio óptico (Nikon, Japón) para obtener la imagen y el número de neuronas supervivientes (mm 2 ) de la corteza isquémica.
Tinción TUNEL
Las secciones de parafina se desparafinaron y deshidrataron con alcohol al 100%, 95%, 80% y 70%. Luego, las secciones se sumergieron en paraformaldehído al 4%, seguido de incubación con tampón de citrato de sodio Triton X-100 al 0,1% durante 20 min y la mezcla de reacción TUNEL durante 1 a 1,5 h. A continuación, las secciones se revelaron mediante una solución de peroxidasa y diaminobencidina (DAB), se contratiñeron con hematoxilina, se deshidrataron con alcohol en gradiente, se permeabilizaron y se sellaron en goma neutra. Las secciones se observaron con un microscopio óptico para contar las células positivas a TUNEL. Las partículas de color amarillo parduzco en el núcleo se definieron como células TUNEL-positivas (células apoptóticas).
Inmunohistoquímica
Las secciones se desparafinaron, hidrataron y recuperaron con antígeno citrato, y la catalasa endógena se bloqueó con H 2 al 3%. O 2 . A cada sección se le añadió suero de cabra (50 μL), se incubó con el anticuerpo primario (50 μL) y se hizo reaccionar con el anticuerpo secundario (50 μL) y estreptavidina marcada con peroxidasa de rábano picante (50 μL). Las secciones se revelaron mediante DAB y se contratiñeron con hematoxilina, a lo que siguió una deshidratación con alcohol en gradiente, permeabilización con xileno y sellado de goma neutra. Las secciones se observaron al microscopio (el citoplasma o el núcleo de los tejidos cerebrales isquémicos eran partículas amarillas o amarillo parduzco). Se midieron y promediaron los valores de DO.
RT-qPCR
Trizol (Invitrogen) realizó la extracción de ARN total de tejidos y células. La expresión de ARNm se analizó mediante qPCR utilizando SYBR Green PCR Mix (Applied Biosystems, CA, EE. UU.) y el sistema de PCR en tiempo real QuantStudio TM 6Flex (Applied Biosystems). Se diseñaron cebadores específicos de transcripción inversa de tallo-bucle y cebadores directo / inverso (BioTNT, Shanghai, China) para analizar los niveles de miARN. La Tabla 1 enumeró todos los cebadores. El 2 - △△ CT Se adoptó un método para calcular los niveles de miARN o ARNm.
Análisis de Western Blot
Las muestras de proteína se separaron mediante electroforesis en gel de poliacrilamida con dodecilsulfato de sodio al 10% y se sometieron a electrotransferencia sobre una membrana de fluoruro de polivinilideno (PVDF). Después de eso, la membrana de PVDF se bloqueó con leche desnatada en polvo al 5% en solución salina que contenía Tris, se probó con los anticuerpos primarios y se volvió a sondar con el anticuerpo secundario durante 2 h. Luego, las bandas de proteína se analizaron con tampón de fosfatasa alcalina que contenía cloruro de tetrazolio nitroazul y fosfato de 5-bromo-4-cloro-3-indolilo y se cuantificaron mediante el software ImageJ (NIH, Bethesda, MD, EE. UU.). Los principales anticuerpos fueron STAT3 (1:1000), Mfn2 (1:1000, Abcam, MA, EE. UU.) Y gliceraldehído-3-fosfato deshidrogenasa (GAPDH) (1:1000, Cell Signaling Technology, Beverly, MA, EE. UU.).
Ensayo génico indicador de luciferasa dual
El STAT3 3'UTR que contiene miR-326-5p de tipo salvaje (WT) o sitios de unión mutante (Mut) se clonó en el vector pmirGLO (Promega, WI, EE.UU.), denominado STAT3-WT y STAT3-Mut. Las células se sembraron en una placa de 24 pocillos a 2 × 10 4 células / pocillo y cotransfectadas con 100 ng de vector de luciferasa STAT3-WT o STAT3-Mut y agomir miR-326-5p 50 nM o agomir NC al 70% de confluencia. La actividad de luciferasa se midió utilizando el sistema de ensayo indicador de luciferasa dual (Promega Corporation, WI, EE. UU.).
Análisis estadístico
Para el análisis de los datos se utilizó el software estadístico SPSS 21.0 (IBM, NY, EE. UU.). Los datos se expresaron como media ± desviación estándar (DE). La diferencia entre dos grupos se analizó mediante la prueba t de Student, mientras que las comparaciones múltiples se analizaron mediante un análisis de varianza unidireccional (ANOVA), seguido de la prueba post hoc de Tukey. P representaba una prueba de dos caras y P <0,05 se consideró estadísticamente significativo.
Resultados
La regulación ascendente de miR-326-5p o la regulación descendente de STAT3 atenúan la apoptosis de las neuronas corticales tratadas con OGD
Cultivadas durante una semana, las neuronas corticales de rata se diferenciaron y maduraron. Microscópicamente, las neuronas mostraban cuerpos neuronales más grandes, citoplasma más transparente, núcleo grande con baja densidad y fuerte índice de refracción. Los cuerpos neuronales eran cónicos o redondos y las dendritas estaban escalonadas y adheridas a las paredes (Fig. 1a, b). NES fue un marcador químico específico de neuronas cerebrales para localizar neuronas con alta especificidad a través de un ensayo de inmunofluorescencia. Se encontró que las neuronas se unían específicamente a NES y mostraban fluorescencia verde (Fig. 1c).
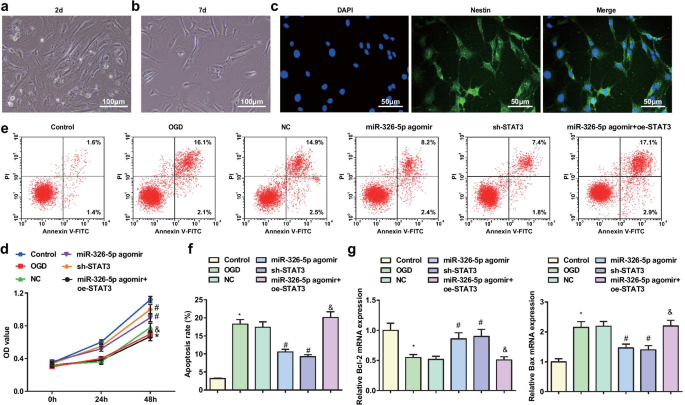
La regulación por aumento de miR-326-5p o la regulación por disminución de STAT3 atenúa la apoptosis de las neuronas corticales tratadas con OGD. un Neuronas cerebrales con cultivo adherente 2-d (barra de escala:100 μm); b Neuronas cerebrales con cultivo adherente de 7 días (barra de escala:100 μm); c Neuronas cerebrales con cultivo adherente de 7 días teñido por ensayo de inmunofluorescencia NES (barra de escala:50 μm); d Viabilidad de neuronas corticales tratadas con OGD mediante ensayo MTT; e Apoptosis de neuronas corticales tratadas con OGD detectadas por citometría de flujo; f Tasa de apoptosis de neuronas corticales tratadas con OGD; g Expresión de ARNm de Bcl-2 y Bax en neuronas corticales tratadas con OGD; * P <0,05 en comparación con el grupo de control; # P <0,05 en comparación con el grupo NC; & P <0.05 en comparación con el grupo agomir miR-326-5p
Se aplicó el ensayo MTT para detectar la viabilidad de las neuronas corticales (Fig. 1d). Los resultados demostraron que la viabilidad de la neurona cortical de rata se vio afectada en el grupo OGD y el grupo miR-326-5p agomir + oe-STAT3 frente al grupo de control y el grupo miR-326-5p agomir, pero mejoró en el grupo miR-326-5p grupo agomir y grupo sh-STAT3 versus grupo NC (todos P <0.05).
La apoptosis de neuronas corticales de rata se determinó mediante citometría de flujo, mientras que la expresión de ARNm de Bcl-2 y Bax mediante RT-qPCR (Fig. 1e-g). Los resultados explicaron que en comparación con el grupo de control y el grupo agomir miR-326-5p, el grupo OGD y el grupo miR-326-5p agomir + oe-STAT3 demostraron un aumento de la tasa de apoptosis, una reducción de la expresión de Bcl-2 y un aumento de la expresión de Bax. (todos P <0,05). Por el contrario, con los grupos NC, se redujo la tasa de apoptosis, se elevó el nivel de Bcl-2 y se suprimió el nivel de Bax en el grupo agomir miR-326-5p y el grupo sh-STAT3 (todos P <0.05).
La regulación ascendente de miR-326-5p o la regulación descendente de STAT3 aumenta el nivel de potencial de la membrana mitocondrial y Mfn2 en las neuronas corticales después de una lesión por OGD
La expresión de Mfn2 en neuronas corticales tratadas con OGD se detectó mediante análisis de transferencia Western (Fig. 2a, b). Con respecto al grupo control y al grupo agomir miR-326-5p, la expresión de la proteína Mfn2 se redujo en el grupo OGD y miR-326-5p agomir + oe-STAT3, mientras que aumentó en el grupo miR-326-5p agomir y sh -Grupo STAT3 versus grupo NC (todos P <0.05).
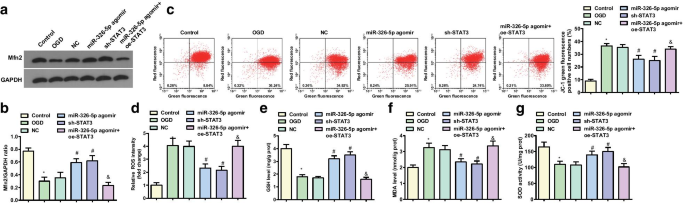
La regulación hacia arriba de miR-326-5p o la regulación hacia abajo de STAT3 aumenta el nivel de potencial de membrana mitocondrial y Mfn2 en neuronas corticales tratadas con OGD. un , b Expresión de la proteína Mfn2 en neuronas corticales tratadas con OGD; c Citometría de flujo JC-1 del nivel de potencial de membrana mitocondrial en neuronas corticales tratadas con OGD; d Contenido de ROS en neuronas corticales tratadas con OGD después de regular al alza miR-326-5p o regular a la baja STAT3; e Actividad de GSH en neuronas corticales tratadas con OGD después de regular al alza miR-326-5p o regular a la baja STAT3; f Contenido de MDA en neuronas corticales tratadas con OGD después de regular al alza miR-326-5p o al regular STAT3; g Actividad de SOD en neuronas después de regular al alza miR-326-5p o regular a la baja STAT3; * P <0,05 en comparación con el grupo de control; # P <0,05 en comparación con el grupo NC; & P <0.05 en comparación con el grupo agomir miR-326-5p
El potencial de membrana mitocondrial detectado por JC-1 encontró que (Fig.2c, d) con respecto al grupo de control y al grupo agomir miR-326-5p, el nivel de potencial de membrana mitocondrial disminuyó en el grupo OGD y el agomir miR-326-5p + Grupo oe-STAT3 (todos P <0,05). En relación con los grupos NC, el nivel de potencial de la membrana mitocondrial aumentó en el grupo agomir miR-326-5p y el grupo sh-STAT3 (todos P <0.05).
Se determinaron los contenidos de ROS y MDA, y las actividades de GSH y SOD en neuronas corticales tratadas con OGD y los resultados indicaron que frente al grupo de control y el grupo de miR-326-5p agomir, el grupo OGD y el miR-326-5p agomir + oe -El grupo STAT3 presentó un aumento en el contenido de ROS y MDA y actividades de GSH y SOD deterioradas (todas P <0,05). En comparación con los grupos NC, el grupo agomir miR-326-5p y el grupo sh-STAT3 mostraron contenidos reducidos de ROS y MDA y actividades reforzadas de GSH y SOD (todas P <0.05) (Fig. 2e – g).
La regulación hacia arriba de miR-326-5p o la regulación hacia abajo de STAT3 impide el daño patológico de las neuronas corticales e inhibe la apoptosis en ratas con IC / RI
Se adoptó la tinción HE para observar el daño patológico de los tejidos cerebrales (Fig. 3a, b), mostrando que en el grupo simulado, las neuronas eran de estructura normal y ordenadas con citoplasma rojo pálido, núcleo azul y nucleolo claro. En el grupo MCAO, el grupo NC y el grupo miR-326-5p agomir + oe-STAT3, a excepción de la necrosis, se observó que las neuronas estaban dispuestas desordenadamente y teñidas más oscuras, con ruptura de la membrana nuclear, desaparición de la estructura celular, cariopyknosis, profunda núcleo teñido y lisis en gran cantidad. En el grupo agomir miR-326-5p y el grupo sh-STAT3, la inflamación de las neuronas se alivió y las neuronas se dispusieron ordenadamente, y las células necróticas se redujeron, lo que indica un mejor estado en relación con el grupo MCAO y el grupo NC.
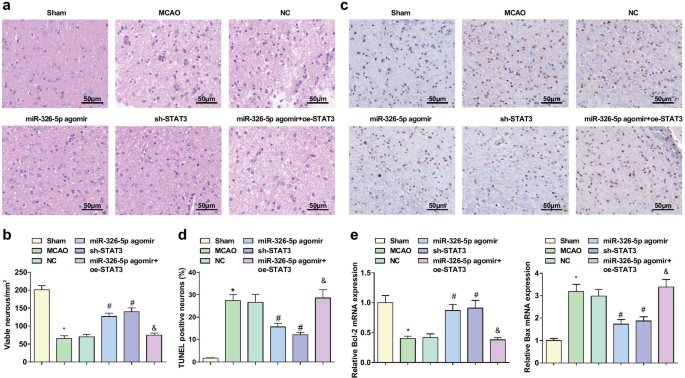
La regulación positiva de miR-326-5p o la regulación negativa de STAT3 impide el daño patológico de las neuronas corticales e inhibe la apoptosis en ratas con CI / RI. un Daño patológico de los tejidos cerebrales en ratas MCAO observado por tinción HE (barra de escala:50 μm); b El número de neuronas intactas de los tejidos cerebrales de rata MCAO después de regular al alza miR-326-5p o regular a la baja STAT3; c Apoptosis neuronal en tejidos de cerebro de rata MCAO detectada por tinción TUNEL (barra de escala:50 μm); d Tasas positivas de TUNEL en tejidos cerebrales de rata MCAO después de regular al alza miR-326-5p o regular a la baja STAT3; e Expresión de ARNm de Bcl-2 y Bax en tejidos de cerebro de rata MCAO después de regular hacia arriba miR-326-5p o regular hacia abajo STAT3; * P <0,05 en comparación con el grupo simulado; # P <0,05 en comparación con el grupo NC; & P <0.05 en comparación con el grupo agomir miR-326-5p
La tinción TUNEL se utilizó para la detección de apoptosis neuronal y RT-qPCR para la expresión de ARNm de Bcl-2 y Bax en tejidos cerebrales. Los resultados mostraron que (Fig. 3c-e) en comparación con el grupo simulado y el grupo agomir miR-326-5p, la tasa TUNEL-positiva aumentó, la expresión de ARNm de Bcl-2 disminuyó y la expresión de ARNm de Bax aumentó en el grupo de MCAO y el grupo miR-326-5p agomir + oe-STAT3 (todos P <0,05). Se mostró una tasa TUNEL positiva más baja y una expresión de ARNm de Bax, y una expresión de ARNm de Bcl-2 más alta en el grupo miR-326-5p agomir y sh-STAT3 que en el grupo NC (todos P <0.05).
La regulación ascendente de miR-326-5p o la regulación descendente de STAT3 aumenta el nivel potencial de la membrana mitocondrial en los tejidos cerebrales de ratas con CI / RI
La detección de la expresión de la proteína Mfn2 en tejido cerebral de rata se realizó mediante inmunohistoquímica, y los resultados mostraron que (Fig.4a, b) se probó una menor expresión de la proteína Mfn2 en el grupo simulado y el grupo agomir miR-326-5p frente al grupo MCAO y el grupo miR-326-5p agomir + grupo oe-STAT3, mientras que se midió una mayor expresión de la proteína Mfn2 en el grupo miR-326-5p agomir y el grupo sh-STAT3 en lugar del grupo NC (todos P <0.05).
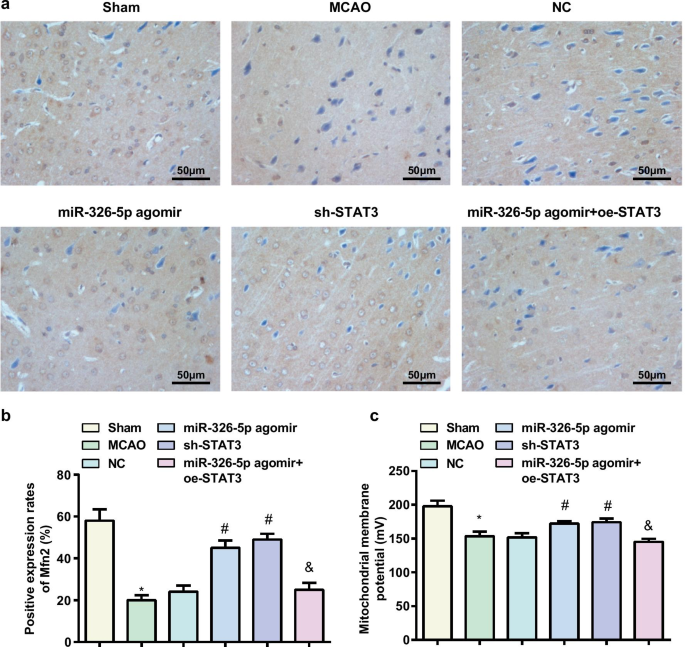
La regulación por aumento de miR-326-5p o la regulación por disminución de STAT3 aumenta el nivel de potencial de la membrana mitocondrial en los tejidos cerebrales en ratas con CI / RI. un Expresión de Mfn2 detectada por inmunohistoquímica en tejido cerebral de rata (barra de escala:50 µm); b Número de células positivas para Mfn2 en tejidos de cerebro de rata MCAO después de regular al alza miR-326-5p o regular a la baja STAT3; c Potenciales transmembrana mitocondrial en neuronas de tejidos cerebrales de rata MCAO después de regular por aumento miR-326-5p o de regulación por disminución de STAT3; * P <0,05 en comparación con el grupo simulado; # P <0,05 en comparación con el grupo NC; & P <0.05 en comparación con el grupo agomir miR-326-5p
La detección del nivel de potencial de membrana mitocondrial descubrió que (Fig. 4c) en comparación con el grupo simulado y el grupo agomir miR-326-5p, el nivel de potencial de membrana mitocondrial disminuyó en el grupo MCAO y el miR-326-5p agomir + oe-STAT3 grupo, mientras que aumentó en el grupo agomir miR-326-5p y el grupo sh-STAT3 en contraste con el grupo NC (todos P <0.05).
miR-326-5p se dirige a STAT3
La expresión de miR-326-5p y STAT3 en neuronas corticales y tejidos cerebrales se detectó mediante RT-qPCR y análisis de transferencia Western. En neuronas corticales in vitro, se sugirió una disminución en la expresión de miR-326-5p y un aumento en la expresión de STAT3 en el grupo OGD en relación con el grupo de control (ambos P <0,05). Con respecto al grupo NC, la expresión de miR-326-5p se elevó y la expresión de STAT3 se redujo en el grupo agomir de miR-326-5p (ambos P <0,05); Expresión de STAT3 disminuida ( P <0,05) en el grupo sh-STAT3. En contraste con el grupo agomir miR-326-5p, la expresión de STAT3 se elevó en el grupo agomir + oe-STAT3 miR-326-5p ( P <0.05) (Fig. 5a – c).
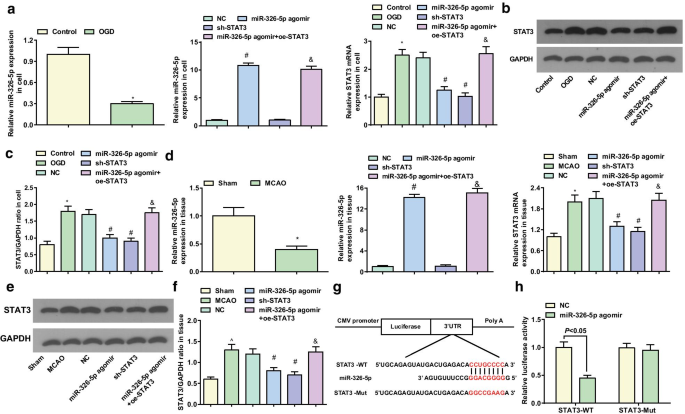
el miR-326-5p apunta a STAT3. un expresión de ARNm de miR-326-5p y STAT3 en neuronas corticales tratadas con OGD in vitro; b , c Expresión de la proteína STAT3 en neuronas corticales tratadas con OGD in vitro; d expresión de ARNm de miR-326-5p y STAT3 en tejidos de cerebro de rata MCAO; e , f Expresión de la proteína STAT3 en tejidos cerebrales de rata MCAO; g Sitio de unión previsto de miR-326-5p y STAT3 mediante software de bioinformática; h La relación de dirección entre miR-326-5p y STAT3 validada por ensayo de gen indicador de luciferasa dual; En la figura a - c , * P <0,05 en comparación con el grupo de control; # P <0,05 en comparación con el grupo NC; & P <0,05 en comparación con el grupo agomir miR-326-5p. En la figura d - f , * P <0,05 en comparación con el grupo simulado; # P <0,05 en comparación con el grupo NC; & P <0.05 en comparación con el grupo agomir miR-326-5p
En los tejidos cerebrales in vivo, en relación con el grupo simulado, aparecieron un miR-326-5p más bajo y un STAT3 más alto en el grupo MCAO (ambos P <0,05). En relación con el grupo NC, la expresión de miR-326-5p aumentó mientras que la expresión de STAT3 disminuyó en el grupo de agomir miR-326-5p (ambos P <0,05). La expresión de STAT3 disminuyó en el grupo sh-STAT3 ( P <0,05). Por el contrario, con el grupo agomir miR-326-5p, la expresión de STAT3 se elevó en el grupo agomir + oe-STAT3 miR-326-5p ( P <0.05) (Fig. 5d – f).
DIANA y el software miRDB encontraron los posibles sitios de dirección de miR-326-5p en STAT3 3'UTR (Fig. 5g). Se construyeron STAT3 3'UTR que contenían el sitio de unión de WT miR-326-5p y el sitio de unión de Mut y se insertaron en el plásmido pmirGLO. La unión de miR-326-5p a STAT3 3'UTR se validó adicionalmente mediante un ensayo indicador de luciferasa. El ensayo indicador de luciferasa reveló que miR-326-5p agomir redujo la actividad luciferasa relativa de STAT3-WT, pero no STAT3-Mut (Fig.5h), lo que implica que STAT3 era un gen diana directo de miR-326-5p.
Discusión
La IC / IR es la principal causa de muerte cerebrovascular [22]. Se indica que los miARN están implicados en CI / RI [23]. Sin embargo, la comprensión integral del mecanismo relacionado con miR-326-5p aún es insuficiente en CI / RI. Por lo tanto, este estudio tiene como objetivo descubrir los roles concretos de miR-326-5p en CI / RI con énfasis en el recíproco combinado de miR-326-5p y STAT3. Productivamente, este estudio ha declarado que la regulación positiva de miR-326-5p atenuó el CI / RI y la expresión de Mfn2 elevada a través de la inhibición de STAT3.
Al principio, se determinó que la expresión de miR-326-5p se reducía en CI / RI in vivo e in vitro. Posteriormente, organizamos una serie de ensayos y descubrimos que la regulación positiva de miR-326-5p mejora la viabilidad neuronal y la función mitocondrial, la expresión de Mfn2 elevada y el estrés oxidativo restringido y la apoptosis. Además, los experimentos in vivo en ratas verificaron aún más las funciones funcionales del miR-326-5p restaurado en CI / RI. De hecho, se ha demostrado que un miR-326 alto se correlaciona con una supervivencia global prolongada de los pacientes con glioblastoma, el tipo común de tumor cerebral [24]. Also, miR-326 expression has been further evidenced to reduce in Parkinson's disease and miR-326 could suppress apoptosis of dopaminergic neurons and reduce inflammatory response [25]. Except for that, the reduction in miR-326-5p is manifested in endothelial progenitor cells in the course of myocardial infarction, and introduction of endothelial progenitor cells overexpressing could promote cardiac function recovery [11]. There has been a study illustrating that up-regulation of miR-326 improves the behavioral function, enhances neuronal viability and depresses neuronal apoptosis in mice with Alzheimer's disease [26]. Furthermore, it is documented that up-regulation of miR-326 by miR-326 mimic could suppress inducible nitric oxide synthase in dopaminergic neurons in Parkinson's disease [27].
In this study, we found that STAT3 was a target gene of miR-326-5p. Supported by an advanced study, it is indicated that STAT3 expression is negatively regulated by miR-326 in human endometrial carcinoma stem cells [28]. In the present study, we measured that STAT3 expression was enhanced in CI/RI. Currently, it has been found that I/R injury and OGD would induce STAT3 expression to increase [29]. Intriguingly, STAT3 mRNA expression trends to an increment in I/R animals [30]. Additionally, STAT3 protein expression is reported to up-regulate in CI/RI [31]. Next, our study revealed that down-regulating STAT3 had the therapeutic effects on CI/RI rats and OGD-treated neurons. As supported by a study, it is concluded that miR-31 induction discourages oxidative stress by inactivation of JAK/STAT3 pathway in ischemic stroke [32]. Also, inhibition of JAK2/STAT3 pathway is beneficial to oxidative stress impairment in CI/RI [31]. Furthermore, knockdown of JAK/STAT3 pathway is identified to enhance cell viability, and reduce oxidative stress and neuron apoptosis in ischemic brain injury [33]. Lately, inhibited JAK/STAT3 signaling pathway is witnessed to relieve myocardial infarction in myocardial I/R injury [34]. As demonstrated, depleted STAT3 undermines neuronal apoptosis in rats with white matter injury [35]. Moreover, the suppression of neuronal apoptosis, and alleviation of cerebral infarct size are attributed to JAK2/STAT3 inhibition in rats with CI/RI [36].
Finally, we focused on the effects of miR-326-5p and STAT3 on Mfn2 expression and discovered that the reduced level Mfn2 in rats with CI/RI and OGD-treated neurons could be elevated by either restoring miR-326-5p or silencing STAT3. As implicated by a previous study, it is documented that Mfn2 is decreased in the lately phase of reperfusion and poorly expressed of Mfn2 exacerbates CI/RI via restrained autophagosome formation and autophagosome and lysosome fusion [18]. Additionally, Mfn2 is implied to reduce even disappear in I/R injury in old hepatocytes [37]. However, few researches have discussed the regulatory mechanism of miR-326-5p and STAT3 for Mfn2.
Conclusión
In general, this study has elucidated the mechanisms that elevated miR-326-5p inhibits neuronal apoptosis, attenuates pathological damage of neurons and increases the expression of Mfn2 via STAT3 downregulation in CI/RI. The study may update the potential mechanism of miR-326-5p and STAT3 in CI/RI. However, more studies are still needed for further development of the molecular mechanism in CI/RI.
Disponibilidad de datos y materiales
No aplica.
Abreviaturas
- miRNAs:
-
MicroRNAs
- STAT3:
-
Signal transducer and activator of transcription-3
- OGD:
-
Oxygen and glucose deprivation
- MCAO:
-
Middle cerebral artery occlusion
- ROS:
-
Especies reactivas de oxígeno
- STAT:
-
Signal transducers and transcriptional activator
- Mfn2:
-
Mitofusin-2
- FITC:
-
Fluorescein isothiocyanate
- NES:
-
Nestin
- NC:
-
Control negativo
- Oe:
-
Sobreexpresión
- RT-qPCR:
-
Reacción en cadena de la polimerasa cuantitativa con transcripción inversa
- OD:
-
Densidad óptica
- PI:
-
Yoduro de propidio
- MDA:
-
Malondialchehyche
- GSH:
-
Glutathione
- SOD:
-
Superóxido dismutasa
- DAB:
-
Diaminobenzidine
- PVDF:
-
Fluoruro de polivinilideno
- WT:
-
Wild-type
- Mut:
-
Mutant
- SD:
-
Standard deviation
- ANOVA:
-
Análisis de varianza unidireccional
Nanomateriales
- Daños y prevención de incendios
- La regulación a la baja del microARN-342-5p o la regulación al alza de Wnt3a inhibe la angiogénesis y mantiene la estabilidad de la placa aterosclerótica en ratones con aterosclerosis
- Preparación de nanopartículas de mPEG-ICA cargadas con ICA y su aplicación en el tratamiento del daño celular H9c2 inducido por LPS
- Compuesto híbrido Nanoestructurado de sílice / oro-celulosa-Amino-POSS mediante el proceso Sol-Gel y sus propiedades
- Eficacia antitumoral y farmacocinética mejoradas de la bufalina mediante liposomas pegilados
- Síntesis fácil de óxido de estaño mesoporoso similar a un agujero de gusano a través del autoensamblaje inducido por evaporación y las propiedades mejoradas de detección de gas
- Diseño y ajuste de la función de trabajo del grafeno mediante tamaño, modificación, defectos y dopaje:un estudio de teoría del primer principio
- Nuevas nanopartículas dirigidas al receptor CD44 y mitocondrial dual para liberación activada por estímulos redox
- Reducción de la conductividad de protones mejorada y la permeabilidad del metanol mediante biomembrana de óxido de grafeno sulfonado con electrolito de alginato de sodio
- Material y propiedades ópticas de los puntos cuánticos de carbono fluorescente fabricados a partir de jugo de limón mediante una reacción hidrotermal
- Propiedades microestructurales, magnéticas y ópticas de las nanopartículas de perovskita manganita La0.67Ca0.33MnO3 sintetizadas mediante el proceso Sol-Gel