


Posibles efectos osteoinductores de las nanopartículas de hidroxiapatita en las células madre mesenquimales mediante la interacción de las células endoteliales
Resumen
La nanohidroxiapatita (nano-HA) ha atraído una atención sustancial en el campo de la medicina regenerativa. Las interacciones entre células endoteliales (CE) y células madre mesenquimales (MSC) son necesarias para la reconstrucción ósea, pero se desconoce la forma en que el nano-HA interactúa en este proceso. En este documento, investigamos la citotoxicidad y los efectos osteoinductivos de las nanopartículas de HA (HANP) en las MSC utilizando un modelo de cocultivo indirecto mediado por CE y resaltamos los mecanismos subyacentes. Se encontró que a una dosis subcitotóxica, los HANP aumentaron la viabilidad y la expresión de los genes de los osteoblastos, así como los nódulos mineralizados y la producción de fosfatasa alcalina de las MSC. Estos fenómenos se basaron en HIF-1α secretado por EC, que desencadenó la cascada de señalización ERK1 / 2. Además, se estableció un modelo matemático de linaje celular en dos etapas para analizar cuantitativamente el impacto de HIF-1α en la diferenciación osteogénica de las MSC. Demostró que HIF-1α ejercía un efecto estimulante dependiente de la dosis sobre la tasa de diferenciación osteogénica de las CMM de hasta 1500 pg / ml, lo que estaba de acuerdo con los resultados anteriores. Nuestros datos implicaron que las interacciones cooperativas entre HANP, CE y MSC probablemente sirvan para estimular la regeneración ósea. Además, el modelo de linaje celular en dos etapas es un sistema in vitro útil para evaluar la influencia potencial de las moléculas efectoras en la ingeniería del tejido óseo.
Introducción
La reconstrucción de defectos óseos causados por traumatismos, malformaciones congénitas o resección quirúrgica plantea un gran desafío para la cirugía ortopédica [1]. La hidroxiapatita (HA), una cerámica bioactiva representativa, se había empleado como sustituto óseo [2]. Sin embargo, las propiedades mecánicas y osteoinductivas indeseables restringen su aplicación clínica [3]. En los últimos años, nano-HA ha demostrado una bioactividad mejor y más óptima y un rendimiento mecánico mejorado debido a sus características biónicas únicas y ha ganado un interés significativo en los campos biomédicos relacionados con la medicina regenerativa [4]. Cuando se implanta nano-HA en defectos óseos, múltiples células involucradas en la reparación ósea estarán expuestas a él. Como tal, es necesario evaluar el comportamiento biológico de nano-HA. Varias líneas de evidencia han demostrado directamente que las nanopartículas de HA (HANP) pueden ser captadas por las células madre mesenquimales derivadas de la gelatina de Wharton (hWJ-MSC) y las células osteoblásticas del cordón umbilical humano, lo que da como resultado una diferenciación osteogénica mejorada [5,6,7]. Dua y col. informó anteriormente de la capacidad de los HANP para promover la integración de cartílago diseñado en cartílago de novo [8]; por el contrario, los HANP inhiben la capacidad angiogénica de las células endoteliales de la vena umbilical humana (HUVEC) [9]. En términos de salud humana, se justifica una comprensión más completa del impacto de los HANP en la regeneración ósea, y las aplicaciones en curso de hueso artificial de nano-ingeniería se suman a la urgencia de tales estudios.
La regeneración ósea va acompañada inevitablemente de la invasión de neovasos. Las CE son el revestimiento celular interno del sistema vascular que sirven para suministrar sangre de forma pasiva y también desempeñan un papel en la inducción, especificación y guía de la regeneración de órganos, así como en el mantenimiento de la homeostasis y el metabolismo [10, 11]. Las CMM son parte del nicho periendotelial y poseen capacidades de autorrenovación y diferenciación múltiple bajo la inducción de microambientes fisiológicos y bioquímicos particulares dentro de sus nichos residentes [12]. Tsai y col. encontraron que las CE pueden secretar endotelina-1 para dirigir las CMM hacia la diferenciación de linaje osteo y condro [13]. Además, Saleh et al. utilizaron análisis de datos de microarrays para identificar proteínas secretadas por HUVEC y vías de señalización de diafonía relacionadas que interactúan con los receptores de membrana de MSC para mejorar la proliferación y la diferenciación osteogénica [14]. En la ingeniería de tejido óseo, los HANP pueden entrar en contacto con los neovasos y ser endocitosados por las CE, lo que ha demostrado alterar la función fisiológica de estas células [9, 15]. Esto también puede influir en las células osteoprogenitoras circundantes y afectar la reparación ósea al alterar la señalización paracrina. Sin embargo, si bien se ha explorado el impacto directo de los HANP en las MSC, todavía existe una falta de una comprensión clara de si los HANP pueden inducir indirectamente la diferenciación osteogénica de las MSC a través de las CE, lo cual es esencial para nuestra comprensión del efecto de los HANP en relación con a la reparación ósea.
En este estudio, en un esfuerzo por obtener más información sobre los efectos biológicos de los HANP en la interacción entre las CE y las MSC, se estableció un modelo de cocultivo indirecto utilizando HUVEC y hWJ-MSC. Mediante la utilización de este sistema, se evaluaron la citotoxicidad y los efectos osteoinductores de los HANP en las hWJ-MSC a través de la señalización paracrina mediada por HUVEC. Para identificar los factores clave que influyen en las interacciones entre células endoteliales y MSC inducidas por HANP, se evaluaron los factores solubles en el sobrenadante de HUVEC que habían sido estimulados con HANP, haciendo hincapié en los mecanismos relacionados tanto a nivel de genes como de proteínas. Los resultados demostraron que el factor inducible por hipoxia (HIF) -1α juega un papel fundamental en estas interacciones.
Para observar y predecir cuantitativamente el impacto de HIF-1α en el proceso de osteogénesis, se estableció un modelo matemático que combina un linaje celular en dos etapas con HIF-1α. Aquí, al analizar los datos empíricos, se utilizó el modelo de linaje celular de dos etapas para predecir el número de MSC y el grado de diferenciación en cualquier momento, según la densidad de siembra celular inicial definida y la concentración de HIF-1α, y esto a su vez puede proporcione sugerencias adecuadas para las condiciones iniciales de cultivo y los tiempos de incubación. Los resultados de este estudio ayudarán a arrojar luz sobre las interacciones entre los sustitutos óseos de base nanométrica y los sistemas biológicos, que pueden servir para promover el desarrollo de biomateriales innovadores para su uso en la medicina regenerativa.
Materiales y métodos
Preparación y caracterización de partículas
Se adquirieron HANP a 20 nm (np20), 20 * 80 nm (np80) y partículas de HA de tamaño micro (m-HAP) con una pureza ≥ 99,0% de Nanjing Emperor Nano Material Company Ltd (Nanjing, China). El tamaño y la forma de las partículas se observaron mediante microscopía electrónica de transmisión (TEM; FEI Tecnai G2 Spirit Bio-Twin, FEI, Hillsboro, OR, EE. UU.) Y microscopía electrónica de barrido (SEM; LEO1530VP, Alemania). El tamaño hidrodinámico y el potencial zeta de las partículas de HA (HAP) se determinaron mediante Zetasizer Nano ZS90 y Mastersizer 3000 (Malvern Instruments, Malvern, Reino Unido).
Preparación y cultivo celular
Todos los protocolos experimentales fueron aprobados por el Comité de Ética de la Universidad Médica de Nanjing. Las HUVEC y las hWJ-MSC se obtuvieron de cordones umbilicales humanos frescos, como se describió anteriormente, después de obtener el consentimiento informado por escrito de los donantes [16, 17]. Brevemente, el cordón umbilical y la vena umbilical se enjuagaron con solución salina tamponada con fosfato (PBS) que contenía penicilina y estreptomicina al 1% (PS; Hyclone, GE Healthcare Life Sciences, Pasching, Austria). Luego, la vena umbilical se llenó con colagenasa I al 0,1% (Sigma, St. Louis, MO, EE. UU.) Y se incubó durante 15 min a 37 ° C. Después de la recolección, las HUVEC se cultivaron en medio EC (ECM) (Sciencell, San Diego, CA, EE. UU.).
Posteriormente, se extrajeron los vasos sanguíneos y se cortó la gelatina de Wharton en 1 mm 2 piezas y luego se colocan en 25 cm 2 matraces de cultivo de tejidos (Corning Incorporated, Corning, NY, EE. UU.). Estas células se incubaron en L-DMEM (GIBCO Life Technology, Grand Island, NY, EE. UU.) Complementado con suero bovino fetal al 10% (GIBCO) y PS al 1%.
Se evaluaron las hWJ-MSC para confirmar el fenotipo utilizando anticuerpos monoclonales contra CD13, CD29, CD34, CD44, CD45, CD51 y CD105 (BD Biosciences, San José, CA, EE. UU.). Las HUVEC se evaluaron mediante el factor von Willebrand (vWF; Shanghai ChangDao Biotech Co, Ltd., Shanghai China). En estos experimentos se utilizaron HUVEC entre los pasajes 3 a 7 y hWJ-MSC entre los pasajes 3 a 5.
A continuación, se diluyeron suspensiones de partículas a 1 mg / ml en PBS en L-DMEM hasta la concentración final. Como se muestra en la Fig. 1, las HUVEC se incubaron en la concentración indicada de HAP durante 18 h. El medio de cultivo se centrifugó a 15.000 rpm a 4 ° C durante 15 min, y los sobrenadantes suplementados con FBS al 10% se utilizaron como medio acondicionado (CM) para hWJ-MSC para realizar los siguientes experimentos. El CM consistió en medio osteogénico suplementado con líquido de inducción osteogénica, que contenía 10 mM de β-glicerofosfato, 50 μg / ml de ácido L-ascórbico-2-fosfato y dexametasona 0,1 μM (Sigma-Aldrich, St Louis, MO, EE. UU.). Además, se usó 2-metoxiestradiol (2-MeOE2) (Selleck Chemicals, Houston, TX, EE. UU.) Como inhibidor específico de HIF-1α. En el grupo 2-MeOE2 (+), las hWJ-MSC se cultivaron con CM de HUVEC, que se pretrataron con 2-MeOE2 5 μM durante 40 minutos antes de la suplementación con HAP. Se utilizó PD98059 como inhibidor específico de MEK. En el grupo PD98059, las hWJ-MSC se cultivaron con CM que contenía PD98059 5 μM.
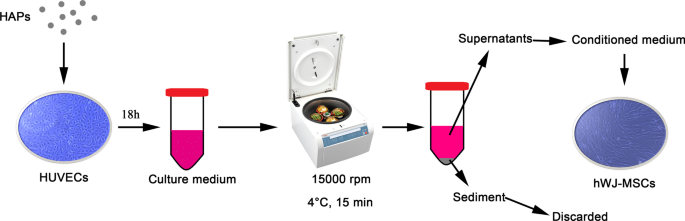
Ilustración de cocultivo indirecto de HAP / hWJ-MSC mediado por HUVEC. Abreviaturas: HAPs partículas de hidroxiapatita, hWJ-MSCs Cordón umbilical humano Células madre mesenquimales derivadas de gelatina de Wharton, HUVEC células endoteliales de la vena umbilical humana
Determinación de la viabilidad y el número de células
La viabilidad celular se evaluó utilizando el kit de ensayo MTS (MTS; Bestbio, Beyotime Biotechnology, Shanghai, China). Las hWJ-MSC se dejaron adherir durante 24 hy luego se cultivaron con CM durante 24 y 72 h. La absorbancia del formazán se evaluó a 490 nm usando un lector de microplacas (SpectraMax M2; Molecular Devices LLC, Sunnyvale, CA, EE. UU.). La absorbancia también se convirtió en números de células utilizando curvas de calibración estándar en las mismas condiciones.
Reacción cuantitativa en cadena de la polimerasa en tiempo real (RT-PCR)
Se usó reactivo TRIzol (Invitrogen, Carlsbad, CA, EE. UU.) Para aislar el ARN total de las células hWJ-MSC incubadas en CM durante el tiempo indicado. El ADN complementario se transcribió a partir de 1,0 µg de ARN usando un kit de síntesis de ADNc PrimeScript First Strand (TaKaRa, Tokio, Japón) en un termociclador T3 (Mastercycler 5333; Eppendorf, Hamburgo, Alemania). Los niveles de expresión de los genes indicados se analizaron utilizando el kit FastStart Universal SYBR Green Master (ROX) (Roche, Basilea, Suiza) en un sistema de amplificación cuantitativa en tiempo real (7900HT Fast; Applied Biosystems, Foster City, CA, EE. UU.). La expresión relativa de ARNm del gen diana se normalizó a GAPDH y luego se determinó usando el método \ (2 ^ {{- \ Delta \ Delta C_ {t}}} \). Las secuencias de cebadores para los genes diana se enumeran en la Tabla 1.
Análisis cuantitativo y tinción con rojo de alizarina S (ARS)
Las hWJ-MSC se cultivaron en placas de 12 pocillos con medio osteogénico durante hasta 14 d, y luego, se observó la mineralización de la matriz extracelular utilizando tinción ARS (Leagene, Leagene Biotechnology, Beijing, China) después de. Brevemente, las muestras se fijaron con alcohol etílico absoluto durante 15 min y luego se tiñeron con ARS al 1% (p / v) (pH, 4,2) a temperatura ambiente durante 5 min. Las células teñidas se lavaron dos veces con agua bidestilada y luego se fotografiaron. Para el análisis cuantitativo del proceso de mineralización, se agregaron a cada pocillo 300 μL de cloruro de cetilpiridinio monohidrato al 10% (p / v) (BOMEI, BOMEI Biotechnology, Hefei, China) y las placas se incubaron durante 30 min. Se transfirió un total de 90 μl de cada muestra a una placa de 96 pocillos y luego se midió la absorbancia a 405 nm por triplicado.
Análisis cuantitativo y tinción de fosfatasa alcalina (ALP)
Después de cultivar las hWJ-MSC en placas de 12 pocillos con medio osteogénico durante hasta 14 días, se realizó la tinción de ALP usando un kit de desarrollo de color de fosfatasa alcalina BCIP / NBT (Beyotime). Brevemente, las hWJ-MSC se fijaron en paraformaldehído al 4%. Luego, las muestras se tiñeron en una mezcla de nitro-azul tetrazolio y 5-bromo-4-cloro-3-indolil-fosfato durante 4 horas y se fotografiaron. Para cuantificar la síntesis de ALP, las células se lisaron en tampón de lisis RIPA helado (Beyotime) durante 30 min. Los lisados celulares se centrifugaron a 12.000 rpm a 4ºC durante 10 min, y los sobrenadantes se sometieron a análisis cuantitativo de ALP usando un kit de ensayo de ALP (Beyotime). La absorbancia se midió por triplicado a 405 nm y se convirtió en actividad ALP utilizando una curva estándar.
Ensayo inmunoabsorbente ligado a enzimas (ELISA)
Las HUVEC se sembraron a 2 × 10 5 células / pocillo. Se recogió CM de HUVEC cultivadas con HAP durante 18 h (archivo adicional 1), y el sobrenadante se sometió a análisis ELISA (Fig. 1). Se utilizó un kit ELISA de HIF-1α humano (Anhui Joyee Biotechnics, Anhui, China) de acuerdo con las instrucciones del fabricante.
Inmunofluorescencia
Se sembraron HUVEC en portaobjetos en placas de 12 pocillos (7,6 x 10 4 células / pocillo). Después de la exposición a CM durante 18 h, las células se fijaron en paraformaldehído al 4% (Biosharp, Beijing, China) y se permeabilizaron con Triton X ‐ 100 (Beyotime) al 0,1% en PBS antes de la incubación con anticuerpo anti-HIF-1α al 1% [EPR16897 ] (ab179483, Abcam, Reino Unido) durante la noche a 4 ℃. Posteriormente, las células se incubaron con IgG anti-conejo de cabra (H + L) conjugado con CoraLite594 al 1% (Proteintech, EE. UU.) En la oscuridad durante 1 h. Luego, los núcleos se tiñeron con DAPI (Beyotime, Shanghai, China), que se añadió a las células y reaccionó durante 30 s. Las muestras se examinaron utilizando un microscopio confocal láser (Olympus, Japón). La intensidad de la fluorescencia se cuantificó utilizando el software de análisis ImageJ v.1.4 (Bethesda, MD, EE. UU.).
Análisis de Western Blot
Las hWJ-MSC se incubaron en CM durante 24 h (quinasa regulada por señal extracelular (ERK) 1/2, p-ERK1 / 2) o en medio osteogénico durante 7 d (factor de transcripción relacionado con runt (RUNX) -2, tipo 1 colágeno / colágeno 1 (COL I)). Luego, las células se lisaron en tampón de lisis RIPA durante 30 min. Los lisados celulares se centrifugaron y los sobrenadantes se almacenaron a -20 ° C para su análisis. Después de SDS-PAGE al 12%, las proteínas se transfirieron a una membrana de difluoruro de polivinilideno (PVDF). Los anticuerpos primarios utilizados fueron anti-p-ERK1 / 2, anti-ERK1 / 2, anti-RUNX-2, anti-COL I y GAPDH (1:1,000, anticuerpos policlonales de conejo; Cell Signaling Technology, Boston, MA, EE. UU. ). Después de eliminar los anticuerpos no unidos, la membrana se incubó con anticuerpos secundarios durante 1 h. La señal en las membranas se detectó utilizando un sistema de imágenes en gel de quimioluminiscencia (LAS4000M; GE Healthcare Biosciences AB, Uppsala, Suecia). La relación de p-ERK a ERK y RUNX-2 / COL I a GAPDH se cuantificó utilizando el software de análisis ImageJ v.1.4 (Bethesda).
Evaluación de la apoptosis celular
Las hWJ-MSC se sembraron a una densidad de 10 5 células por pocillo en placas de 6 pocillos. Las células adherentes se trataron con las concentraciones indicadas de HIF-1α durante el tiempo indicado. A continuación, se recogieron las células y se marcaron con FITC-Anexina V y PI (Fcmacs, Nanjing, China) durante 15 minutos en la oscuridad. Todas las muestras se analizaron con un citómetro de flujo FACScan (BD Bioscience, San José, CA, EE. UU.). Los datos se analizaron con FlowJo v10 (BD Biosciences).
Modelo de linaje celular de dos etapas
Teniendo en cuenta el enorme potencial de los HAP y la dificultad de analizar el sistema de cocultivo, se requirió un modelo matemático que pudiera proporcionar un análisis cuantitativo y se requirió una predicción confiable para comprender el papel de HIF-1α en la diferenciación osteogénica de hWJ- MSC.
Las hWJ-MSC se cultivaron con 0, 300, 500, 1000, 1500, 2000, 3000 y 4000 pg / ml de HIF-1α, así como con fluido de inducción osteogénica. Después de ajustar los datos a estas concentraciones, usamos las concentraciones de HIF-1α (producido por HUVEC) en los grupos control, m-HAP, np80 y np20 (240, 300, 325 y 375 pg / mL, respectivamente) para pruebe las ecuaciones de ajuste utilizando MATLAB (MathWorks, Natick, MA, EE. UU.). Para simplificar el modelo, consideramos que sus patrones de crecimiento similares a las diferentes concentraciones de HIF-1α son idénticos. Se empleó el grado de diferenciación promedio para ajustar la ecuación de tiempo de grado de diferenciación. La tasa de proliferación, la tasa de apoptosis y los grados de diferenciación osteogénica de las hWJ-MSC en los diferentes grupos se detectaron en momentos definidos.
Un modelo simplificado de linaje celular de dos etapas, que era similar a un modelo de linaje celular de múltiples etapas [18, 19], se estableció de acuerdo con los datos experimentales. C 0 y C 1 representan el número de celda de las hWJ-MSC y las celdas terminales, respectivamente. C 0 y C 1 se rigen por:
$$ \ left \ {\ begin {recolectado} \ frac {{{\ text {d}} C_ {0}}} {{{\ text {d}} t}} =\ left [{\ frac {{K - C_ {0} - C_ {1}}} {K} p - (p - 1)} \ right] \ upsilon_ {0} C_ {0} \ hfill \\ \ frac {{{\ text {d}} C_ {1}}} {{{\ text {d}} t}} =\ left ({2 - \ frac {{K - C_ {0} - C_ {1}}} {K} p - p} \ derecha) \ upsilon_ {0} C_ {0} - AC_ {1} \ hfill \\ \ end {collect} \ right. $$Aquí, p , afectado por HIF-1α y el tiempo, representa la probabilidad de replicación de las hWJ-MSC. En consecuencia, d =1 - p es la tasa de diferenciación que se puede obtener ajustando los datos estimados y experimentales del número de células. El parámetro v 0 cuantifica la rapidez con que las células se dividen en cada etapa del linaje (en particular, v =Ln2 / c , donde c es la duración de un ciclo celular). La tasa de apoptosis de las células terminales está simbolizada por A . Para simplificar, descuidamos que la tasa de apoptosis variará ligeramente con el tiempo y, por lo tanto, A =4.5% es una constante. K denota la capacidad ambiental porque las células no pueden experimentar una proliferación ilimitada [20]. HIF-1α aumenta la tasa de diferenciación de las hWJ-MSC, lo que lleva a una tasa de diferenciación modelada por:
$$ \ begin {alineado} d &=\ frac {{d_ {0}}} {{1 + (r * H) ^ {m}}} \\ p &=1 - d \\ \ end {alineado} $$Aquí, H representa la concentración de HIF-1α; d 0 indica la tasa de diferenciación a 0 pg / ml de HIF-1α; r representa la intensidad de la regulación (en este documento, que representa la intensidad de la regulación de HIF-1α en las MSC); ym corresponde al coeficiente de Hill [21], scilicet la relación entre la tasa de diferenciación de MSC y la concentración de HIF-1α.
Análisis estadístico
Todos los datos que cumplen con los requisitos de normalidad y homocedasticidad se expresan como la media ± desviación estándar (DE) de tres o más experimentos independientes. Se utilizó el software SPSS 24.0 (SPSS Inc., Chicago, EE. UU.) Para realizar los análisis estadísticos mediante ANOVA de una vía o ANONA de dos vías. A P valor <0,05 se consideró estadísticamente significativo. El análisis estadístico se presenta utilizando GraphPad Prism 5 (GraphPad Software, La Jolla, CA).
Resultados
Caracterización de HAP
Como se muestra en la Fig. 2, los HAP se prepararon con un tamaño y forma particulares. Los diámetros del np20 con forma casi esférica eran de 20 nm en promedio, y el np80 tenía forma de varilla con una longitud promedio de 80 nm y un ancho de 20 nm. Los m-HAP también eran de forma casi esférica y de aproximadamente 12 μm de diámetro. Todas las partículas tenían una carga superficial negativa en L-DMEM. Se ha sugerido que los valores negativos del potencial zeta tienen un efecto favorable significativo sobre la unión y proliferación de las células óseas, así como la unión ósea directa y la formación de hueso nuevo [22, 23]. Las partículas, que se observaron en L-DMEM, tienen tendencia a agregarse en sistemas acuosos. También se evaluó su tamaño hidrodinámico, que también podría ser un factor importante que afecte sus comportamientos biológicos.

Caracterización de HAP. Micrografías TEM de a np20 y b np80 y micrografía SEM de c m-HAP. d Caracterización de HAP ( n =6). Abreviaturas: TEM microscopía electrónica de transmisión, SEM microscopía electrónica de barrido, HA hidroxiapatita, m-HAP partículas de HAP de tamaño micro
Toxicidad indirecta de los HAP hacia las hWJ-MSC
Para evaluar la toxicidad indirecta de los HAP en hWJ-MSC, se midió la viabilidad celular mediante ensayos MTS. CM con 50 µg / mL de HANPs podría estimular significativamente la viabilidad de hWJ-MSC después de 24 hy 72 hy especialmente a las 24 h. Sin embargo, CM con 100 µg / mL de HANP disminuyó la viabilidad celular en un 15-20% en comparación con el control después de 72 h. Además, CM con 25 µg / mL np20, pero no np80, estimuló la viabilidad celular después de 24 h. Estos fenómenos confirmaron que los HANP de 50 µg / ml eran una concentración subcitotóxica que se utilizó en todos los experimentos posteriores (Fig. 3).
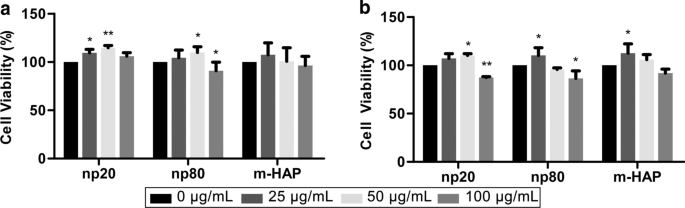
Toxicidad indirecta de HAP hacia hWJ-MSC. Se midió la viabilidad de hWJ-MSC cultivadas indirectamente con HAP para a 24 y b 72 h. * P <0,05; ** P <0,01 frente al control. El grupo de control consistió en células incubadas en CM sin tratamiento con HAP, y la viabilidad celular se normalizó como porcentaje del control. Abreviaturas: HAPs partículas de hidroxiapatita, m-HAP partículas de HAP de tamaño micro, hWJ-MSC Cordón umbilical humano Células madre mesenquimales derivadas de la gelatina de Wharton
Efecto osteoinductivo indirecto de HAP en hWJ-MSC
Para identificar los efectos osteoinductores indirectos de los HANP en las hWJ-MSC, se evaluó la expresión de genes osteogénicos mediante análisis cuantitativo de RT-PCR. El nivel de transcripción para el factor de transcripción 2 relacionado con runt (RUNX-2) en los grupos HANP, especialmente np20, exhibió un aumento notable desde el Día 7 hasta el Día 21 (Fig. 4a). La expresión génica del colágeno de tipo I (Col I) en el grupo np20 demostró una mejora desde el día 7 hasta el día 21, mientras que el grupo np80 demostró un aumento sostenido desde el día 7 hasta el día 14 (Fig. 4b). El ARNm de osteocalcina (OCN) estaba claramente regulado positivamente en los grupos HANP el día 14, lo que indica una tasa acelerada de osteogénesis (Fig. 4c). Los niveles de ARNm de fosfatasa alcalina (ALP) obviamente aumentaron en los grupos HANP (Fig. 4d), lo que indica la diferenciación osteogénica de hWJ-MSC. Sin embargo, el grupo de m-HAP presentó cambios limitados con respecto a los niveles de estos tres genes osteogénicos en comparación con el grupo de control (Fig. 4). Además, la expresión de marcadores de pluripotencia, NANOG, OCT3 / 4 y SOX2, disminuyó en los grupos HANP en comparación con el control (Fig.4e-g), lo que implica que las hWJ-MSC en los grupos HANP se habían diferenciado, especialmente en el grupo np20. Se obtuvieron resultados similares mediante análisis de transferencia Western (Fig. 5c, d), lo que indica que el grupo np 20 podría mejorar indirectamente la expresión de RUNX-2 y COL I en hWJ-MSC.
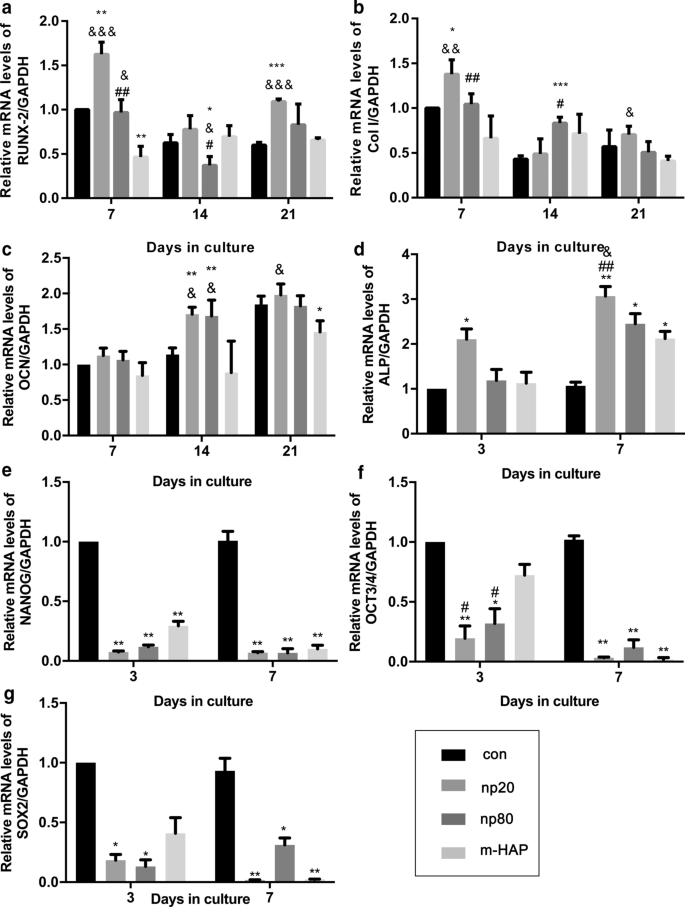
Efectos indirectos de los HAP sobre la expresión de genes relacionados con la diferenciación osteogénica. un RUNX-2, b Col I, c OCN, d ALP, e NANOG, f OCT3 / 4 y g Niveles del gen SOX2 en hWJ-MSC cultivadas con CM durante el tiempo indicado. * P <0,05; ** P <0,01; *** P <0,001 frente al grupo de control; y P <0,05; && P <0,01; &&& P <0,001 frente al grupo m-HAP; # P <0,05; ## P <0,01 frente al grupo np20. Las células incubadas en medio osteogénico sin tratamiento con HAP se utilizaron como grupo de control. Abreviaturas: HAPs partículas de hidroxiapatita, m-HAP partículas de HAP de tamaño micro, hWJ-MSC Cordón umbilical humano Células madre mesenquimales derivadas de la jalea de Wharton, RUNX-2 factor de transcripción 2 relacionado con runt, Col I colágeno tipo I, OCN osteocalcina, ALP fosfatasa alcalina, SOX 2 HMG-box 2 relacionado con SRY
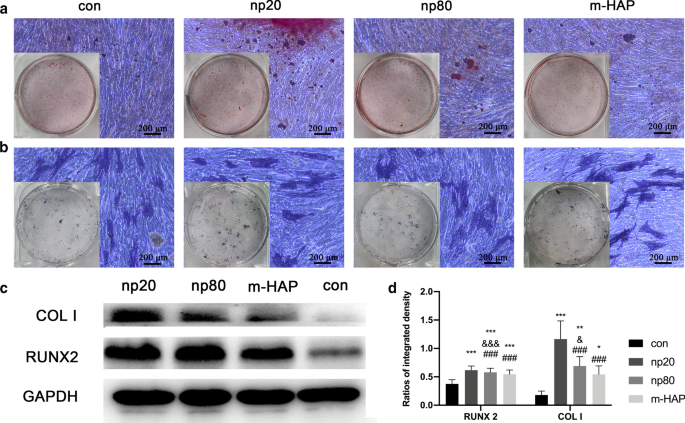
Efecto indirecto de los HAP sobre la deposición extracelular de calcio y la actividad de la ALP. Las hWJ-MSC se incubaron en medio osteogénico durante 14 d. un A continuación, se visualizó la deposición de calcio extracelular mediante tinción con ARS; b La actividad ALP de las hWJ-MSC se evaluó mediante tinción ALP, barras de escala:200 µm. c El análisis de transferencia Western indicó la expresión de RUNX-2 y COL I de hWJ-MSC en medio osteogénico el día 7. d Mediciones densitométricas de RUNX-2 y COL I de la pieza ( c ). Las células incubadas en medio osteogénico sin tratamiento con HAP se utilizaron como grupo de control. * P <0,05; ** P <0,01; *** P <0,001 frente al grupo de control; y P <0,05; &&& P <0,001 frente al grupo m-HAP; ### P <0,001 frente al grupo np20. Abreviaturas: HAPs partículas de hidroxiapatita, m-HAP partículas de HAP de tamaño micro, hWJ-MSC Cordón umbilical humano Células madre mesenquimales derivadas de la jalea de Wharton, ALP fosfatasa alcalina
Para observar visualmente el efecto osteoinductivo indirecto de los HANP sobre las hWJ-MSC, las células se incubaron con el medio osteogénico indicado durante 14 d, seguido de tinción con ARS y ALP. Como se muestra en la Fig. 5a, b, se observó un mayor número de nódulos mineralizados y una mayor actividad ALP de las hWJ-MSC en los grupos HANP en comparación con los grupos m-HAP y control. Además, m-HAP, similar al control, demostró efectos limitados sobre la diferenciación osteogénica de las hWJ-MSC.
Señalización ERK1 / 2 activada por HAP en hWJ-MSC cocultivado indirectamente con HUVEC
Para investigar los efectos sobre la función paracrina de las HUVEC por las HAP, se utilizaron ensayos de inmunofluorescencia y ELISA para identificar la posible proteína que promueve la diferenciación osteogénica de las hWJ-MSC. Como se muestra en la Fig. 6a-c, la producción intracelular y extracelular de HIF-1α fue facilitada significativamente por HANP (especialmente np20), mientras que hubo un efecto limitado de m-HAP en su producción.
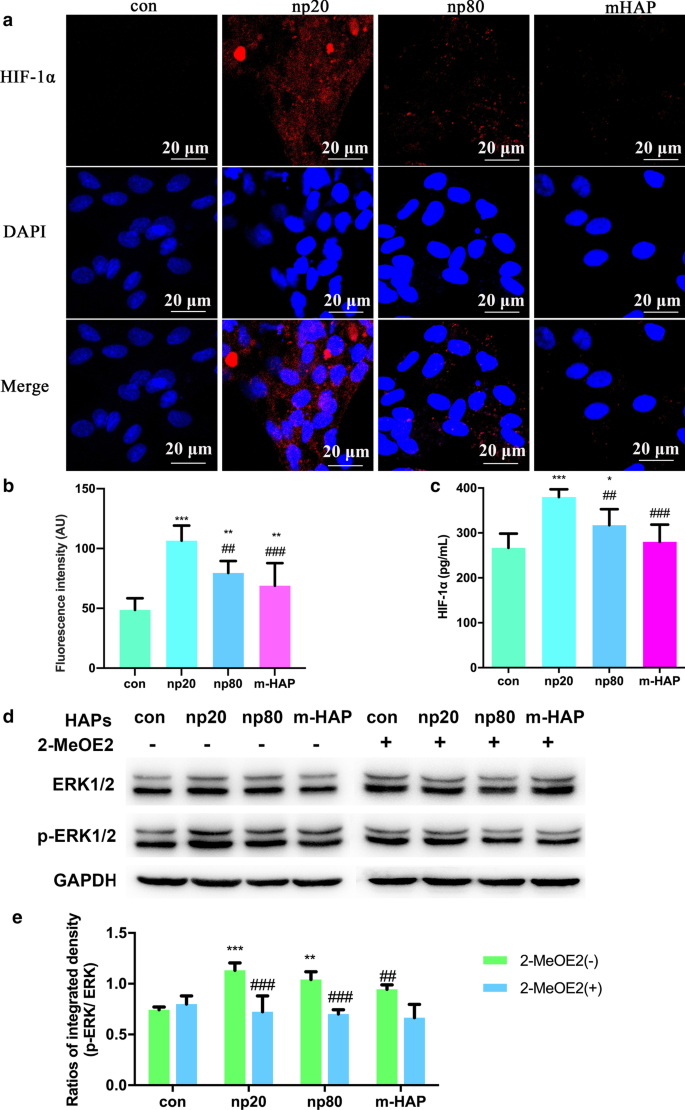
Los HAP activaron la señalización de ERK1 / 2 en hWJ-MSC cocultivadas indirectamente con HUVEC. un Inmunofluorescencia de HIF-1α realizada en HUVEC tratadas con / sin HAP durante 18 h. b La intensidad de fluorescencia de HIF-1α de la parte ( a ). c La concentración extracelular de HIF-1α en el medio de cultivo de HUVEC tratadas con / sin HAP durante 18 h se midió mediante ELISA. Barras de escala:20 μm. * P <0,05; ** P <0,01; *** P <0,001 frente al control; ## P <0,01; ### P <0,001 frente al grupo np20. Las células sin tratamiento con HAP se utilizaron como grupo de control. Las hWJ-MSC se trataron con CM durante 24 h. d Análisis de transferencia de Western que indica la activación de quinasas clave en las vías ERK1 / 2. e Mediciones densitométricas de p-ERK1 / 2 de la parte ( b ). ** P <0,01; *** P <0,001 frente al control; ## P <0.01, ### P <0,001 frente al grupo 2-MeOE2 (-). Las células incubadas en CM sin tratamiento con HAP se utilizaron como grupo de control. Abreviaturas: HAPs partículas de hidroxiapatita, m-HAP partículas de HAP de tamaño micro, hWJ-MSC Cordón umbilical humano Células madre mesenquimales derivadas de gelatina de Wharton, HUVEC células endoteliales de la vena umbilical humana, ERK quinasa regulada por señales extracelulares, HIF-1α factor 1α inducible por hipoxia
Para comprender con mayor precisión la vía de señalización de diferenciación de las hWJ-MSC activadas por HIF-1α, examinamos los reguladores clave de la vía ERK1 / 2 mediante análisis de transferencia Western. Como se muestra en la Fig. 6d, e, mientras que los niveles de proteína de ERK1 / 2 total permanecieron inalterados, los niveles de p-ERK1 / 2 aumentaron claramente en las hWJ-MSC cultivadas con HANP, y esto fue especialmente cierto en el grupo np20. Sin embargo, m-HAP tuvo poco efecto sobre los niveles de p-ERK1 / 2 en las hWJ-MSC, similar a su efecto sobre la producción de HIF-1α en las HUVEC. Es importante destacar que los niveles aumentados de p-ERK1 / 2 en las hWJ-MSC activadas por HIF-1α podrían ser bloqueados por 2-MeOE2, un inhibidor específico de HIF-1α, lo que indica que HIF-1α funcionaba corriente arriba de la vía de señalización ERK1 / 2. en hWJ-MSC.
Diferenciación osteogénica promovida por HIF-1α de hWJ-MSC a través de la vía ERK1 / 2
Para determinar si HIF-1α era necesario para la estimulación observada de la diferenciación osteogénica de hWJ-MSC, se aplicó un inhibidor específico de HIF-1α (2-MeOE2) a estos cultivos celulares. Como se muestra en la Fig.7, la deposición de matriz mineralizada y la actividad ALP de las hWJ-MSC del grupo tratado con 2-MeOE2 (+) cultivadas en medio osteogénico se debilitaron, lo que indica que HIF-1α era indispensable para la diferenciación osteogénica de las hWJ- MSC. Basándonos en estos resultados, exploramos aún más el papel de la vía ERK1 / 2 en la diferenciación osteogénica de las hWJ-MSC activadas por HIF-1α. La deposición de matriz mineralizada y la actividad ALP en las hWJ-MSC cultivadas con medio osteogénico se suprimieron después de la administración de PD98059, un inhibidor específico de MEK.
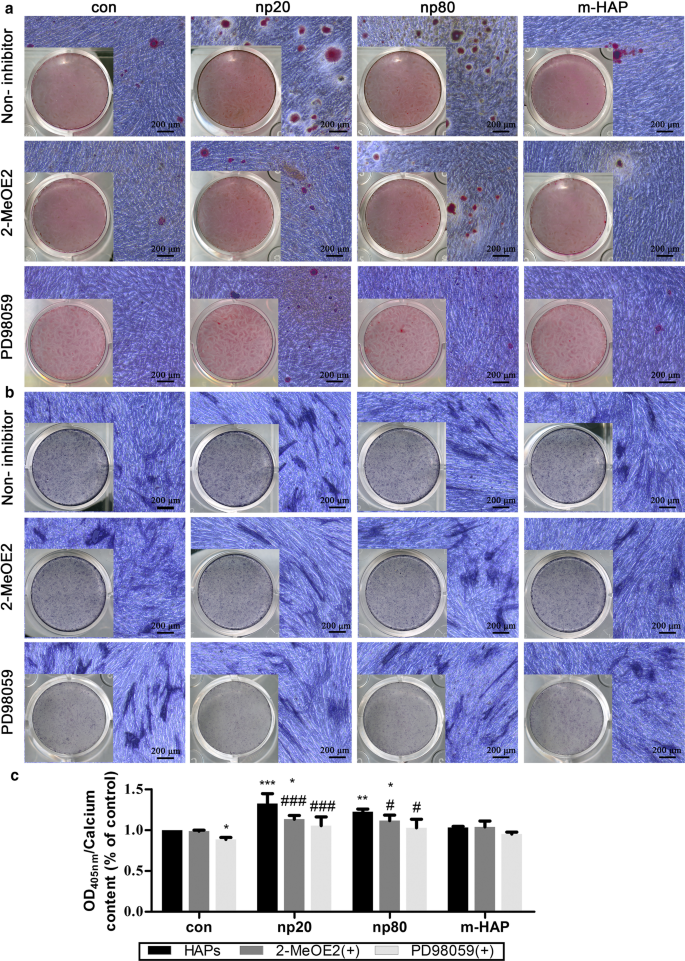
HIF-1α promovió la diferenciación osteogénica de hWJ-MSC a través de la vía ERK1 / 2. Las hWJ-MSC se incubaron en medio osteogénico con o sin PD98059 durante 14 d. un La deposición de calcio extracelular se visualizó mediante tinción con ARS. b La actividad ALP de hWJ-MSC se evaluó mediante tinción ALP. Barras de escala:200 μm. c Análisis cuantitativo de la matriz de calcio extracelular. * P <0,05; ** P <0,01; *** P <0,001 frente al control; # P <0,05; ### P < 0.001 versus the np20 group. Cells incubated in osteogenic medium without HAPs and PD98059 treatment were used as the control group. Abbreviations:HAPs hydroxyapatite particles, m-HAP micro-sized HAP particles, hWJ-MSCs human umbilical cord Wharton’s jelly-derived mesenchymal stem cells, HUVECs human umbilical vein endothelial cells, ERK extracellular signal-regulated kinase, HIF-1α hypoxia-inducible factor 1α, ALP alkaline phosphatase
Two-Stage Cell-Lineage Models
To quantitatively reveal the intrinsic connection between the concentration of HIF and osteogenic differentiation of hWJ-MSCs, 0–4000 pg/mL of HIF-1α was used to treat eight groups of hWJ-MSCs. A two-stage cell-lineage mathematical model was used to analyze the proliferation, apoptosis, and osteogenic differentiation rates of these hWJ-MSCs treated with different concentrations of HIF-1α. As shown in Fig. 8a, fitting data were employed to obtain the simulated formula (\(\frac{d}{{d_{0} }} =\frac{1}{{0.14H^{2} - 0.43H + 1}}\)) and curve (blue curve), showing that the differentiation rate first increased and then decreased with the increase in HIF concentration. More specifically, the differentiation rate reached a peak at 1500 pg/mL HIF-1α.

Two-stage cell-lineage models. hWJ-MSCs were incubated in a defined concentration of HIF-1α for the indicated times. un Relative differentiation rates of hWJ-MSCs at different concentrations of HIF-1α, b relative ALP activity (differentiation degrees of hWJ-MSCs) and relative osteoblast cells on different culture days. c Three-dimensional surface of differentiation degree evolving with time and HIF-1α. Cell number with an initial seeding density of 1500 cells per well (in a 96-well plate), as well as d 0, e 375, and f 1500 pg/mL HIF-1α. Abbreviations:hWJ-MSCs human umbilical cord Wharton’s jelly-derived mesenchymal stem cells, HIF-1α hypoxia-inducible factor 1α, ALP alkaline phosphatase
According to our study, the concentrations of HIF-1α produced by HUVECs in the control, m-HAP, np80, and np20 groups were 240, 300, 325, and 375 pg/mL, respectively. The black square represents the differentiation rate promoted by 240, 300, 325, and 375 pg/mL HIF-1α, and this matches well with the simulated curve. In addition, the differentiation degrees of the hWJ-MSCs (relative ALP activity) treated with different concentrations of HIF-1α increased similarly increased with time. Therefore, in order to simplify the model, we consider their growth patterns to be similar or even identical. We found that the increase in ALP activity from Day 0 to Day 7 was proportional to the number of osteoblasts, and osteoblast cells reached their peak at the platform period. Therefore, after fitting the ALP activity with the relative osteoblasts cell number curve (osteoblasts cell number / maximum osteoblasts cell number), the maximum of ALP activity was predicted and relative ALP activity (ALP activity/ maximum of ALP activity), representing the differentiation degree, was acquired (Fig. 8b). Combining these two studies, the three-dimensional surface of the differentiation degree evolving with time and HIF-1α was obtained (Fig. 8c).
In order to estimate the optimal culture time, it was necessary to simulate cell numbers. After elucidating the differentiation rate under different concentrations of HIF-1α, we simulated the size of each population. The simulation utilized an initial seeding density of 1500 cells per well (in a 96-well plate) using different concentrations of HIF-1α. As shown in Fig. 8d–f, the experimental data (black square) match well with the simulated total cell numbers (blue curve), which is a sum of the number of hWJ-MSCs and osteoblasts, and this supports the ability of this model to predict the number of hWJ-MSCs and osteoblasts at any time point. Moreover, the osteoblast cell number approached the platform period at 21, 18, and 15 days with 0, 375, and 1500 pg/mL HIF-1α, respectively. This model provides the optimum culture time for guiding tissue engineering, and it also provides direct evidence that HIF-1α accelerates the osteogenic differentiation of hWJ-MSCs.
Discusión
With recent advances in nanobiomaterials, nano-based artificial bone substitutes have been an area of intense investigation. The accumulating evidence suggests that there are complex interactions between cells and nanobiomaterials due to their capacity to penetrate cell membranes and increase internal retention times [24, 25]. A previous study revealed that collagen/alginate nanofilms can adsorb onto the MSC membrane to activate intracellular signaling cascades and promote osteogenic differentiation [26]. Elegant experiments by Wu and his colleagues clearly demonstrated that TiO2 nanotubes can improve vascularization and osteogenic differentiation by facilitating paracrine effects and cell junctions via EC-MSC interactions [27]. For the purpose of developing excellent candidates for bone tissue engineering, it is necessary to clarify the direct crosstalk between nano-based bone substitutes and cells implicated in bone repair as well as their indirect interactions. However, our current understanding of this is still limited. In the present study, we utilized an indirect co-culture model to further elucidate the biological effects of HANPs on MSCs in regard to the indirect interactions mediated by ECs.
Cytotoxicity is a primary issue for assessing the biocompatibility of any nanobiomaterial. Although our previous study found that HANPs did not directly influence the viability or apoptosis of hWJ-MSCs, they may still exert different impacts via the mediation of other cells [28]. Thus, it was necessary to evaluate the cytotoxic effects of HANPs on hWJ-MSCs mediated by HUVECs. Interestingly, after incubation in CM for 24 h and 72 h, hWJ-MSC viability was maintained and even elevated in the 0–50 µg/mL HANP groups, especially in the np20 group, indicating the existence of effector molecules in the CM. When the concentration of HANPs reached 100 µg/mL, they became cytotoxic to the hWJ-MSCs. However, 0–100 µg/mL m-HAP had no influence on hWJ-MSC viability (Fig. 3). Jiang y col. have shown that engineered nanoparticles of a particular size can have distinct endocytic routes and kinetics associated with altered downstream signaling involved in regulating target cell functions [29]. In our previous study, we showed that np20 and np80 were endocytosed by HUVECs, and this was followed by morphologic changes and the appearance of large vacuoles, indicating the activated state of the HUVECs. Additionally, np20, with their faster uptake speed and increased accumulation, might result in a stronger activation of HUVECs, possibly resulting in increased hWJ-MSC viability via paracrine signaling. Conversely, few m-HAPs can be endocytosed by HUVECs, and this might account for their limited influence on the metabolism of hWJ-MSCs [9].
To further explore the potential osteoinductive effect of activated HUVECs, a subcytotoxic dose of 50 µg/mL HAPs was used in subsequent studies. The CM collected from the activated HUVECs promoted extracellular calcium deposition, ALP activity, and osteogenic proteins expression in hWJ-MSCs, as well as the mRNA expression of osteogenic genes (Figs. 4, 5). Runx2, an essential transcription factor involved in specifying the osteoblast lineage [30], showed a substantial enhancement in the np20 group, indicating a strong osteoinductive effect on hWJ-MSCs (Fig. 4a and Fig. 5c, d). The np20 group demonstrated a 1.5-fold improvement in COLI expression at Day 7 (Figs. 4b, 5c, d) and a double increase at Day 14, which implied the presence of additional differentiated osteoblasts in the HANP-treated groups (Fig. 4b) [30]. OCN is a mature stage bone marker [31], and this gene showed a significant increase in the HANP groups at Day 14 (Fig. 4c), indicating that np20 and np80 can accelerate bone maturation compared to m-HAP. ALP is an early marker of osteoblast differentiation, and it obviously increased with culture time in each group, especially the np20 group, revealing that additional transformation occurred from MSCs to osteoblasts (Fig. 4d). Pluripotency markers, NANOG, OCT3/4, and SOX2 imply the capacity for differentiation [32]. As shown in Fig. 4e–g, the decreased expression in the genes of the HANP groups implied that most of the hWJ-MSCs in HAP groups had transformed into osteoblasts.
Our data demonstrated that the endocytosis of HANPs by HUVECs was associated with an improved osteogenic differentiation of hWJ-MSCs. However, the cause of this outcome is currently unclear. In terms of the paracrine function of HUVECs, we focused on soluble differentiation-inducing proteins in the supernatant of activated HUVECs. HIF-1α signaling is essential in coupling ossification and angiogenesis during bone regeneration [33, 34]. Heikal et al. reported that injured ECs secrete more HIF-1α even under normoxia conditions [35]. It has also been shown that exposure to HANPs inhibits the angiogenic ability of HUVECs [9]. Thus, we measured the concentrations of HIF-1α in the CM, and the results showed that the HIF-1α content increased in the HANP treatment groups compared to the m-HAP and control groups (Fig. 6a). To identify the role of HIF-1α in the osteogenic differentiation of hWJ-MSCs, we used 2-MeOE2, which is a specific HIF-1α inhibitor, was used. The decreased concentration of HIF-1α paralleled the impaired mineralized matrix deposition and ALP activity in these hWJ-MSCs, indicating that HANPs can promote the HIF-1α production of HUVECs to facilitate the osteogenesis of hWJ-MSCs (Fig. 7).
To properly apply HANPs for use in bone tissue engineering, it is necessary to gain further insights into the mechanisms by which HANPs promote the osteogenic differentiation of hWJ-MSCs mediated by HUVECs. The ERK1/2 pathway is downstream of HIF-1α [36] and is fundamental to the differentiation of MSCs [37]. In this work, the concentrations of HIF-1α in the CM coincide well with the p-ERK1/2 levels in the hWJ-MSCs (Fig. 6b, c). When 2-MeOE2 was applied, the p-ERK1/2 expression in the hWJ-MSCs failed to be activated, indicating that HIF-1α functioned upstream of ERK1/2 signaling. To directly address the role of ERK1/2 signaling in the osteogenic differentiation of hWJ-MSCs, PD98059, a specific MEK inhibitor, was used. The suppression of ERK1/2 signaling resulted in the lowest osteogenic differentiation of hWJ-MSCs. One possible reason for this occurrence is that the ERK1/2 pathway plays a key role in both HIF-1α signaling and in the apoptosis and proliferation signaling pathways, which could be responsible for the observed changes in osteogenic differentiation in these cells [38, 39]. Additionally, this could also be related to the presence of vascular endothelial growth factor (VEGF). VEGF is one of the downstream effectors of HIF-1α signaling [33], and it can also promote the osteogenic differentiation of MSCs via activation of the ERK1/2 pathway [37]. Our previous study found that np20 induced the production of VEGF in HUVECs [9]; therefore, it is possible that the suppression of the ERK1/2 pathway may result in inhibition of VEGF, which would lead to the decreased osteogenic differentiation of hWJ-MSCs. According to the available experimental results, we can summarize as follows. HANPs are able to more optimally process better direct [5] and indirect osteoinductive effects than m-HAPs. Compared to autogenous bone grafts and bone allografts, there is an extensive source of HANP and without secondary damage and potential immunogenicity. However, compared to m-HAPs, HANPs can suppress the angiogenic ability of HUVECs [9] and exhibit slight cytotoxicity in both a time- and dose-dependent fashion.
Recently, growing evidence has demonstrated the importance of HIF-1α in the bone regeneration. However, few studies have been able to quantitatively predict the MSC differentiation rate under specific initial conditions, such as the HIF-1α concentration. Taking cell proliferation, apoptosis, and osteogenic differentiation into account, we present a mathematical model that combines a two-stage cell lineage with HIF-1α that is highly correlated with our experimental data. By fitting the differentiation rate of hWJ-MSCs in 0–4000 pg/mL HIF-1α, we acquired the equations for describing the differentiation rate, HIF-1α concentration, and time. As shown in Fig. 8d, this model can depict the cell number map under different HIF-1α concentrations, so that it is possible to explore the intrinsic dynamics of the two-stage system [40]. Additionally, this model mathematically validates the effect of HIF-1α on the osteogenic differentiation of hWJ-MSCs. Moreover, based on a multi-stage cell-lineage model and logistic model, our model is sufficiently stable to enable long-term predictions without falling into the trap of population unlimited explosion [41].
By using the existing experimental data, both the cell number and differentiation rate can be predicted with a defined initial cell seeding density and HIF-1α concentration. As such, the optimum incubation time is also obtained. Consequently, we can predict the optimum concentration of HIF-1α and determine the most optimal time for osteogenesis, which is important for efficient tissue engineering. A two-stage cell-lineage model is applicable for predicting the proliferation and differentiation of stem cells, which have two cell lineages. On this basis, the model founded on the initial conditions and existing experimental data can be established to identify the optimum culture conditions in vitro, which will assist in optimizing bone repair in vivo.
Conclusión
In this study, we explored the specific biological effects of HANPs on hWJ-MSCs mediated by HUVECs. Compared to m-HAPs, both np20 and np80 showed slight cytotoxicity in both a time- and dose-dependent fashion. Importantly, the size of the HANPs appeared to have no significant impact on this cytotoxicity. Our data also showed that HANPs, especially np20, were capable of facilitating HUVECs to secrete increased levels of HIF-1α, which directly correlated with the enhanced osteogenic differentiation of hWJ-MSCs via the activation of the ERK1/2 pathway (Fig. 9). More remarkably, the results from the two-stage cell-lineage model suggested that HIF-1α exerted a dose-dependent stimulatory effect on the osteogenic differentiation rate of hWJ-MSCs. Additionally, the optimum concentration of HIF-1α and incubation time were estimated based on the initial conditions using an in vitro model, which could be invaluable in the future for tissue engineering applications. Collectively, these observations provide evidence that HANPs may improve bone regeneration by modulating cell–cell interactions.

A schematic illustration of the possible mechanisms
Disponibilidad de datos y materiales
The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
Abreviaturas
- HA:
-
Hydroxyapatite
- HANPs:
-
HA nanoparticles
- hWJ-MSCs:
-
Human umbilical cord Wharton’s jelly-derived mesenchymal stem cells
- HUVECs:
-
Human umbilical vein endothelial cell
- m-HAP:
-
Micro-sized HAP particles
- PBS:
-
Solución salina tamponada con fosfato
- ECM:
-
EC medium
- 2-MeOE2:
-
2-Methoxyestradiol
- ELISA:
-
Ensayo inmunoabsorbente ligado a enzimas
- RUNX-2:
-
Runt-related transcription factor 2
- Col I:
-
Type I collagen
- OCN:
-
Osteocalcin
- ALP:
-
Alkaline phosphatase
- SOX 2:
-
SRY-related HMG-box 2
- ERK:
-
Extracellular signal-related kinases
- VEGF:
-
Factor de crecimiento endotelial vascular
Nanomateriales
- Baterías especiales
- Célula solar
- Análisis de actina y organización de la adhesión focal en células U2OS en nanoestructuras poliméricas
- Las nanotecnologías desactivan las células madre cancerosas
- Estudio in vitro de la influencia de nanopartículas de Au en las líneas celulares HT29 y SPEV
- Efectos de interacción en el ensamblaje de nanopartículas magnéticas
- Dirigirse a las células endoteliales con nanopartículas multifuncionales de GaN / Fe
- Efectos sinérgicos de nanopartículas de Ag / BiV1-xMoxO4 con actividad fotocatalítica mejorada
- Promoción del crecimiento celular SH-SY5Y mediante nanopartículas de oro modificadas con 6-mercaptopurina y un péptido penetrante de neuronas
- Nanocomposites a base de óxido de grafeno decorados con nanopartículas de plata como agente antibacteriano
- Efectos del fullereno C60 sobre la interacción del difenil-N- (tricloroacetil) amidofosfato con el ADN in silico y su actividad citotóxica contra la línea celular leucémica humana in vitro