


F-actina regula la diferenciación osteoblástica de células madre mesenquimales en nanotubos de TiO2 a través de MKL1 y YAP / TAZ
Resumen
El titanio y las aleaciones de titanio se utilizan ampliamente en implantes ortopédicos. La modificación de la nanotopografía proporciona una nueva estrategia para mejorar la osteointegración de sustratos de titanio. La polimerización de actina filamentosa (F-actina), como estructura de carga mecánica, generalmente se considera involucrada en la migración celular, endocitosis, división celular y mantenimiento de la forma celular. Queda por dilucidar si la F-actina está involucrada y cómo funciona en la diferenciación osteogénica inducida por nanotubos de las células madre mesenquimales (MSC). En este estudio, fabricamos TiO 2 nanotubos en la superficie de un sustrato de titanio por oxidación anódica y caracterizaron sus características mediante microscopía electrónica de barrido (SEM), análisis de dispersión de energía de rayos X (EDS) y microscopía de fuerza atómica (AFM). Se realizaron tinciones con fosfatasa alcalina (ALP), Western blot, qRT-PCR y tinción por inmunofluorescencia para explorar el potencial osteogénico, el nivel de F-actina y la expresión de MKL1 y YAP / TAZ. Nuestros resultados mostraron que el diámetro interno y la rugosidad del TiO 2 Los nanotubos aumentaron con el aumento del voltaje de oxidación anódica de 30 a 70 V, mientras que su altura fue de 2 μm de manera constante. Además, cuanto mayor sea el diámetro del tubo, más fuerte será la capacidad del TiO 2 nanotubos para promover la diferenciación osteogénica de las CMM. La inhibición de la polimerización de F-actina por Cyto D inhibió la diferenciación osteogénica de las MSC, así como la expresión de proteínas contenidas en complejos de adhesión focal como la vinculina (VCL) y la quinasa de adhesión focal (FAK). Por el contrario, después del tratamiento con Jasp, la polimerización de F-actina mejoró la expresión de RhoA y los factores de transcripción YAP / TAZ. Con base en estos datos, llegamos a la conclusión de que TiO 2 Los nanotubos facilitaron la diferenciación osteogénica de las CMM, y esta capacidad se mejoró con el diámetro creciente de los nanotubos dentro de un cierto rango (30-70 V). F-actina medió este proceso a través de MKL1 y YAP / TAZ.
Introducción
El titanio y las aleaciones de titanio, debido a su excelente biocompatibilidad, resistencia a la corrosión y propiedades mecánicas, se utilizan ampliamente en aplicaciones clínicas como reemplazos totales de articulaciones e implantes dentales [1, 2, 3]. Sin embargo, todavía quedan muchos desafíos por resolver, incluido el aflojamiento aséptico y la infección [4, 5]. En los últimos años se han llevado a cabo una serie de estudios encaminados a mejorar la osteointegración y las propiedades antibacterianas. Por ejemplo, MoS 2 / El recubrimiento PDA-RGD en implantes de titanio no solo puede promover la integración de un implante de titanio con el hueso huésped, sino que también inhibe el crecimiento bacteriano con una alta eficiencia [6]. Además, la topografía de la superficie ha atraído cada vez más atención, y la modificación topográfica se diferencia de las modificaciones químicas solo cambiando la estructura a micro y nanoescala. La estimulación de señales químicas en las células es inestable y citotóxica. En comparación, las señales físicas seguras y controlables pueden evitar algunos efectos secundarios causados por moléculas químicas. Por tanto, la modificación topográfica de la superficie de los implantes y la regulación de la osteointegración a través de la estructura topográfica pueden proporcionar una nueva forma de solucionar el problema clínico de la mala osteointegración tras la implantación de prótesis.
En el campo de la ingeniería de tejido óseo y la regeneración ósea, la interacción entre la morfología y las células se considera una estrategia de gestión prometedora para el control preciso de la función y diferenciación de las células de la semilla. Al mismo tiempo, el propio hueso tiene una elegante jerarquía dentro del rango de nanómetros y micrones [7]. Por lo tanto, la morfología de la superficie puede proporcionar un nicho similar, que puede imitar la estructura ósea natural y promover la diferenciación osteogénica de las células madre mesenquimales en la superficie del hueso huésped y el implante. Las morfologías de la superficie pueden constar de muchas estructuras diferentes, incluidos nanotubos, nanocables, nanoporos, etc. En particular, las matrices de nanotubos han atraído un gran interés en muchos campos en los últimos años debido a sus características superficiales únicas, como una alta relación superficie-volumen, plasticidad biológica y alta capacidad de adsorción. Por ejemplo, un nuevo estudio muestra que los nanotubos de nitruro de boro (BNNT) constituyen un material sensible al gas que puede usarse como sensor de gas para monitorear el funcionamiento de los transformadores detectando la composición y el contenido de los gases disueltos en el aceite [8]. En biomedicina, la topografía de la superficie también puede dirigir los comportamientos celulares, incluida la migración, adhesión, proliferación y diferenciación celular. Los últimos estudios revelan que la topografía a nanoescala puede hacer que las células madre mesenquimales (CMM) se diferencien en osteoblastos para reforzar la osteointegración temprana [9,10,11,12]. Incluso se ha informado de que la modificación combinada de la superficie a micro y nanoescala puede hacer que las CMM se diferencien en células de músculo liso contráctiles [13]. Sin embargo, los mecanismos moleculares de cómo la topografía de la superficie dirige el destino de las células aún no se han dilucidado, lo cual es importante para la evaluación y el diseño de la seguridad de los materiales.
La (F) -actina filamentosa, también llamada microfilamento, es uno de los tres componentes principales del citoesqueleto en las células eucariotas. Está compuesto por polímeros de (G) -actina globular, modificada por muchas otras proteínas. F-actina tiene polaridad estructural debido al hecho de que todas las subunidades del microfilamento apuntan hacia el mismo extremo. El extremo con púas se dirige a un monómero adyacente diferente, mientras que el extremo puntiagudo posee una subunidad de actina con el sitio de unión de ATP expuesto. Es decir, el ATP está involucrado en el proceso de transformación entre G-actina y F-actina. Este proceso se encuentra en un equilibrio dinámico, con polimerización y despolimerización que ocurren simultáneamente, también conocido como cinta de correr, que a menudo se observa en lamelipodios y filopodios [14]. Por lo tanto, es obvio que la dinámica de la actina juega un papel importante en funciones celulares como la migración celular, la división celular y el mantenimiento de la forma celular. Sin embargo, la F-actina no solo actúa como una estructura física que soporta la carga mecánica, sino que también participa en otros comportamientos biológicos como la transducción de señales y la expresión génica. La evidencia acumulada demuestra que la F-actina puede convertir señales físicas en señales químicas al interactuar con otras proteínas [15,16,17,18]. Por ejemplo, la reconstrucción biomecánica y geométrica promueve la apoptosis de las células tumorales al evitar la superposición de la polimerización del monómero de actina a F-actina [15]. Los pulsos de ultrasonido mejoran la osteogénesis de las células madre mesenquimales humanas al inhibir la despolimerización de la F-actina [16]. Nuestro estudio anterior también mostró que la tensión mecánica aumenta la estabilidad de la F-actina [17]. Debido a que la estructura hueca de los nanotubos proporciona menos sitios de adhesión para las células, el reordenamiento del citoesqueleto es inevitable para mantener el equilibrio biomecánico. En consecuencia, definitivamente tenemos razones para creer que es probable que la F-actina medie en la diferenciación celular inducida por la nanotopografía.
En este estudio, fabricamos TiO 2 nanotubos, modificaron su topografía mediante oxidación anódica y exploraron su capacidad para promover la diferenciación osteogénica de las CMM. A continuación, investigamos si la F-actina desempeña un papel fundamental en la mecanotransducción. La citocalasina D (Cyto D), que se une competitivamente al extremo con púas de la F-actina para evitar que la G-actina se incorpore al filamento, se usó para inhibir la polimerización de la F-actina, y se usó jasplakinolida (Jasp) para mejorar la estabilización de la F-actina. montaje de actina. Además, también queríamos dilucidar cómo funciona la F-actina para cambiar las señales físicas en señales bioquímicas. Basándonos en los resultados de nuestro estudio anterior, planteamos la hipótesis de que la vía MAPK puede estar involucrada en este proceso [17]. Los factores de transcripción como la proteína asociada a Yes (YAP) / coactivador transcripcional con motivo de unión a PDZ (TAZ) y MKL1, que se consideran mecanosensores y mecanotransductores, también fueron el tema de nuestro estudio para analizar cómo la F-actina influye en las células madre destino, porque algunos estudios en otras áreas implicaron que estaban relacionados con F-actina [19,20,21]. En general, esperamos aclarar el papel de la F-actina en el proceso de diferenciación de células madre inducida por nanotubos, de modo que oriente el diseño de materiales y la evaluación de la bioseguridad de los implantes modificados por nanotubos.
Materiales y métodos
Fabricación de TiO 2 Nanotubos
Las rodajas de titanio puro (99,9% de pureza, 2 mm de espesor; Shengshida, Hebei, China), utilizadas como sustrato, se pulieron con papel de lija de carburo de silicio de grano N ° 400 y 1500. A continuación, las muestras se lavaron secuencialmente con acetona, alcohol anhidro y agua desionizada en una máquina de limpieza ultrasónica y finalmente se secaron a temperatura ambiente durante 3 h. Para fabricar la nanotopografía, las muestras pretratadas se fijaron como ánodo, mientras que se utilizó una pieza de platino como contracátodo en una solución acuosa de electrolitos de NH 4 0,15 M F y glicol al 90% durante 1 h. El voltaje de anodización fue un voltaje constante de 30, 40, 50, 60 o 70 V. Después de la oxidación anódica, cada muestra se enjuagó con agua desionizada durante 30 min y se lavó con alcohol anhidro en una máquina de limpieza ultrasónica durante 15 min. Finalmente, todas las muestras se esterilizaron en autoclave a 120 ° C durante 1 h y luego se humedecieron con medio de cultivo antes de su uso.
El mecanismo de reacción de la fabricación de nanotubos no está claro, y la teoría dominante actual es la teoría de la disolución mejorada en el campo. La formación de matrices de nanotubos es el resultado del equilibrio dinámico bajo la acción de la oxidación del campo, la disolución del campo y la disolución química (Fig. 1b). El proceso de anodización se puede describir de la siguiente manera:en el primer paso, se forma una capa de barrera de óxido en la interfaz electrolito-metal:
$$ {\ mathrm {Ti}} ^ {4 +} + 2 {\ mathrm {H}} _ 2 \ mathrm {O} \ to {\ mathrm {Ti} \ mathrm {O}} _ 2 + 4 {\ mathrm { H}} ^ {+} $$ (1)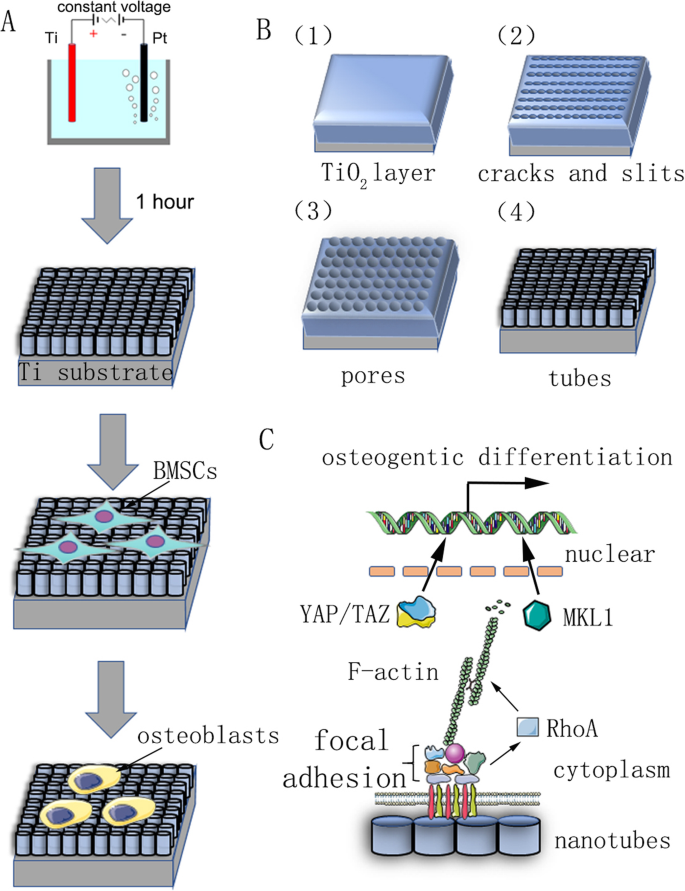
Un cuadro resumen de nuestro estudio. un Diagrama de flujo que muestra la oxidación anódica y la inducción de la diferenciación celular. b El sustrato de titanio pretratado se fijó como ánodo en una solución acuosa de electrolito con NH 4 0,15 M F y glicol al 90% a voltaje constante durante 1 h. Se predijo que los nanotubos autoensamblados se formarían uniformemente. Su mecanismo de reacción se describe en materiales y métodos. c Diagrama esquemático del mecanismo de diferenciación osteogénica de células madre inducida por nanotubos
Luego aparecen grietas y rendijas estrechas en la superficie debido a la disolución de la capa de óxido mejorada en el campo. Difusión de F - iones en estas grietas y rendijas mejora la velocidad de disolución. Las grietas se agrandan y se conectan con las grietas vecinas. Finalmente, la tasa de formación y la tasa de disolución de la capa de óxido de titanio alcanzan un equilibrio dinámico y los nanotubos ya no crecen:
$$ {\ mathrm {TiO}} _ 2 + 6 {\ mathrm {F}} ^ {-} + 4 {\ mathrm {H}} ^ {+} \ to {\ left [{\ mathrm {TiF}} _ 6 \ right]} ^ {2 -} + 2 {\ mathrm {H}} _ 2 \ mathrm {O} $$ (2)Caracterización de superficies
Las muestras fabricadas con diferentes voltajes (30, 40, 50, 60 o 70 V) se enjuagaron con etanol y agua desionizada durante 15 min y luego se secaron a temperatura ambiente. Se utilizó microscopía electrónica de barrido (SEM450, FEI Nova Nano SEM; Thermo Fisher Scientific, Waltham, MA, EE. UU.) Para caracterizar la estructura de la superficie y para medir el diámetro interno y la altura de los nanotubos después de recubrir las muestras con una fina capa de oro. Mientras tanto, se realizó un análisis de dispersión de energía de rayos X (EDS) para analizar la composición elemental de los nanotubos. Se utilizó microscopía de fuerza atómica (AFM, NanoManVS, Bruker Nano Surfaces, Bruker MicroCT, Kontich, Bélgica) para investigar la morfología de la superficie y la rugosidad de la superficie de las muestras. Se seleccionaron tres áreas diferentes de cada muestra y las mediciones se repitieron tres veces.
Cultivo celular
Se compraron ratas macho Sprague-Dawley (SD) de cuatro semanas de edad en el centro de animales experimentales del Noveno Hospital Popular de Shanghai (Shanghai, China). Se aislaron asépticamente células madre mesenquimales de médula ósea de rata (BMSC) de fémures y tibias. Las BMSC se purificaron y expandieron aún más en medio esencial mínimo α (α-MEM; Hyclone, Logan, UT, EE. UU.) Que contenía suero bovino fetal (FBS) al 10% (v / v) (Gibco / Life Technologies, Carlsbad, CA, EE. UU. ), 100 mg / ml de estreptomicina (Gibco) y 100 U / ml de penicilina (Gibco), y se incuban a 37 ° C en una atmósfera humidificada que consta de 95% de aire y 5% de CO 2 . El medio de cultivo se renovó cada 2 días y las células se tripsinizaron y subcultivaron al 80% de confluencia. Todas las células utilizadas en este estudio estaban entre los pases 3 y 5. El medio de inducción osteogénica estaba compuesto por medio de crecimiento suplementado con dexametasona 100 nM, β-glicerofosfato 10 mM y ácido ascórbico 50 mM (Sigma-Aldrich, St Louis, MO, EE. UU.) .
Ensayo de proliferación celular
El TiO 2 Se cortaron rodajas de titanio modificado con nanotubos en formas circulares y se colocaron en pocillos de una placa de cultivo celular de 24 pocillos. Las BMSC entre los pases 3-5 se cultivaron en TiO 2 nanotubos con una densidad de 3 × 10 4 células / disco en medio de crecimiento o medio osteogénico. Después de 2 días de cultivo celular, citocalasina D (Cyto D, Sigma-Aldrich) y jasplakinolide (Jasp, Sigma-Aldrich), utilizados para interrumpir la polimerización de F-actina, se agregaron al medio todos los días durante 3 días. La concentración final y el tiempo de trabajo de Cyto D y Jasp fueron 5 μM, 1 hy 2 μM, 3 h respectivamente. El medio de cultivo se renovó después de la incubación con los reactivos. La viabilidad y la proliferación celular se evaluaron usando el ensayo Cell Counting Kit-8 (CCK8) (Dojindo, Kumamoto, Japón) 12 h después del tratamiento con Cyto D o Jasp. Las células se incubaron con una solución de CCK8 al 10% (v / v) durante 2 h en una incubadora de células a 37 ° C bajo 5% de CO 2 . Luego, transferimos 100 μL de la mezcla de reacción a los pocillos de una placa de 96 pocillos y se midió la absorbancia (DO) del producto de tinte de formazán en el cultivo a 450 nm utilizando un espectrofotómetro Multiscan UV-visible (Safire2; TECAN, Mannedorf, Suiza ). Además, también contamos el número aproximado de células utilizando un contador de células automatizado (AMQAX1000, Life Technologies). Antes del recuento de células, las BMSC se separaron enzimáticamente del TiO 2 nanotubos y teñidos con azul tripán (Sigma-Aldrich).
Tinción de fosfatasa alcalina y análisis de actividad ALP
Se sembraron BMSC en cinco TiO 2 diferentes rodajas de titanio modificado con nanotubos (30, 40, 50, 60 o 70 V) con una densidad de 3 × 10 4 por pocillo y cultivado en medio osteogénico. Los reactivos se agregaron como se describió anteriormente. Después de 7 días de incubación, BMSC cultivadas en TiO 2 Los nanotubos se lavaron tres veces con PBS, se fijaron con paraformaldehído al 4% y se incubaron en una solución de trabajo de fosfatasa alcalina (ALP) de un kit de ALP de acuerdo con las instrucciones del fabricante (Hongqiao, Shanghai, China). Los resultados se observaron bajo un microscopio estereoscópico después de lavarlos con PBS.
Para el análisis de la actividad de ALP, las células se lisaron primero con tampón RIPA sin inhibidores de proteasa y fosfatasa, y luego se analizaron los lisados centrifugados utilizando un kit de ensayo de ALP (Beyotime Institute of Biotechnology, Jiangsu, China) siguiendo el protocolo proporcionado. Finalmente, la actividad se normalizó a la concentración de proteína del lisado correspondiente.
Inmunocitoquímica
Después de 3 días de tratamiento con Cyto D y Jasp, las BMSC se fijaron con paraformaldehído al 4% durante 20 min a temperatura ambiente y luego se lavaron tres veces con PBS. Las células se permeabilizaron con Triton-X 100 al 0,3% durante 30 min, se lavaron tres veces con PBS y se tiñeron con faloidina conjugada con rodamina durante 1 ha temperatura ambiente en la oscuridad. Luego, las células se enjuagaron con PBS y se contratiñeron con DAPI (Instituto de Biotecnología Beyotime) durante 10 minutos a temperatura ambiente. Después de tres lavados adicionales con PBS, las muestras se fijaron en un portaobjetos de vidrio y se observaron mediante microscopía confocal.
Western Blot
Para evaluar la expresión de proteínas, BMSC cultivadas en TiO 2 Los nanotubos se recolectaron con tripsina (Gibco). Las células se lavaron tres veces con PBS y se lisaron con tampón RIPA suplementado con un cóctel inhibidor de proteasa y fosfatasa durante 30 min en hielo. El lisado se recogió mediante centrifugación a 12.000 x g durante 15 min a 4 ° C. La concentración de proteínas totales en el sobrenadante se midió utilizando un kit de ensayo de proteínas de ácido bicinconínico (BCA) (Beyotime) de acuerdo con las instrucciones del fabricante. Se añadió tampón de carga a las muestras de proteína como se indicó anteriormente, que luego se hirvieron a 95 ° C durante 15 min. Para el análisis de transferencia Western, se cargaron 10 μl de la preparación de proteína en un gel SDS-PAGE al 12,5% (EpiZyme Inc., Cambridge, MA, EE. UU.) Y se sometieron a electroforesis a 120 V durante 1 h, luego se transfirieron eléctricamente a un polivinilidenodifluoruro (PVDF) a 250 mA durante 2 h. A continuación, las membranas se bloquearon con leche en polvo desnatada al 5-10% en TBST durante 1 h en un agitador a temperatura ambiente y se incubaron con anticuerpo primario diluido en tampón de dilución (Beyotime) a 4 ° C durante la noche. A continuación, se añadió a las membranas anticuerpo secundario conjugado con fluorescencia diluido en tampón de dilución después de lavar tres veces con TBST durante 5 min, y luego se incubaron las membranas a temperatura ambiente durante 1 h en la oscuridad. Las bandas de proteínas se detectaron mediante un sistema de imágenes de fluorescencia infrarroja de dos colores (Odyssey, LiCor Biosciences, Lincoln, NE, EE. UU.). En particular, si las bandas de la proteína de referencia interna estaban unificadas, la membrana se quitaba y se volvía a analizar con otro anticuerpo primario, seguido del mismo proceso. Usamos el anticuerpo GAPDH como nuestra proteína de referencia interna para normalizar la expresión de la proteína, y los otros anticuerpos primarios utilizados en este estudio fueron anti-vinculina (dilución 1:1000, Abcam, Cambridge, MA, EE. UU.), Anti-FAK (dilución 1:1000 , Tecnología de señalización celular, Danvers, MA, EE. UU.), Anti-Runx2 (dilución 1:1000, tecnología de señalización celular), anti-RhoA (dilución 1:1000, tecnología de señalización celular), anti-F-actina (dilución 1:500 , Abcam), anti-Osx (dilución 1:500, Abcam) y anti-pYAP (dilución 1:1000, tecnología de señalización celular). Los anticuerpos secundarios fueron IgG H&L anti-ratón de cabra (IRDye® 680RD, dilución 1:5000, Abcam) e IgG H&L anti-conejo de cabra (IRDye® 680RD, dilución 1:5000, Abcam).
PCR cuantitativa en tiempo real
Se llevó a cabo una PCR cuantitativa en tiempo real el día 7 para evaluar la expresión génica del factor de transcripción 2 relacionado con runt (Runx2), Osterix (Osx), Alp, osteocalcina (OCN), RhoA, YAP, TAZ, vinculina (VCL), quinasa de adhesión focal (FAK) y leucemia megacarioblástica 1 (MKL1) en células cultivadas en medio osteogénico en TiO 2 nanotubos. El ARN total se extrajo de las células usando un kit de ARN total (R6812-01HP, Omega Bio-Tek Inc., Norcross, GA, EE. UU.). La concentración y pureza de las muestras de ARN se determinaron mediante densidad óptica a una longitud de onda de 260 y solo se analizaron las muestras que presentaban relaciones A260 / 280 y relaciones A260 / 230 superiores a 1,8. Las muestras de ARN se sometieron a transcripción inversa en ADNc utilizando un kit de síntesis de ADNc qScript (Takara, Shiga, Japón) de acuerdo con las instrucciones del fabricante. La PCR cuantitativa en tiempo real se realizó con SYBR® Premix Ex Taq ™ (Takara) utilizando un sistema de PCR en tiempo real QuantStudio 6 Flex (Life Technologies). GAPDH, un gen doméstico, se utilizó como referencia interna. Los datos se analizaron mediante la comparación Ct (2 -ΔΔCt ) y expresado como cambios de veces en comparación con el control. Las secuencias de los cebadores utilizados se enumeran en la Tabla 1.
Análisis estadístico
Todos los datos son representativos de al menos tres experimentos independientes que utilizan muestras por triplicado a menos que se indique lo contrario. Los datos se expresan como media ± desviación estándar (DE). Las diferencias entre los grupos se evaluaron mediante un análisis de varianza unidireccional seguido de la prueba post-hoc de Student-Newman-Keuls o la t de Student. prueba. P los valores <0,05 se consideraron estadísticamente significativos.
Resultados
Caracterización de superficies
Para fabricar la nanotopografía, TiO 2 Los nanotubos se formaron sobre un sustrato de titanio puro utilizando un equipo de oxidación anódica a diferentes voltajes constantes (30, 40, 50, 60 y 70 V) durante 1 h (Fig. 1). Se observó una matriz distribuida uniformemente de nanotubos autoensamblados mediante microscopía electrónica de barrido (SEM). Las vistas lateral y superior de los nanotubos se muestran en la Fig. 2a, b. La altura de los nanotubos en todas las muestras de este estudio fue de aproximadamente 2 μm, mientras que los diámetros internos de los nanotubos fueron de aproximadamente 74 nm (30 V), 92 nm (40 V), 112 nm (50 V), 128 nm (60 V) y 148 nm (70 V) (figura 2c). Esto indicó que la altura de los nanotubos estaba relacionada con el tiempo de oxidación anódica y el diámetro interno estaba relacionado con el voltaje de oxidación anódica. A continuación, se llevó a cabo un análisis de dispersión de energía de rayos X (EDS) para analizar la composición elemental de los nanotubos. Esto mostró que los nanotubos constan de solo dos elementos, O y Ti (Fig. 2d, e). Se utilizó microscopía de fuerza atómica (AFM) para detectar las estructuras nanotubulares y medir la desviación media aritmética del perfil de nanotubos como la rugosidad de la superficie (Ra) (Fig. 2f). Los datos mostraron que la rugosidad de la superficie de los nanotubos aumentaba a medida que aumentaba el diámetro (es decir, el voltaje de oxidación anódica) (Fig. 2g).
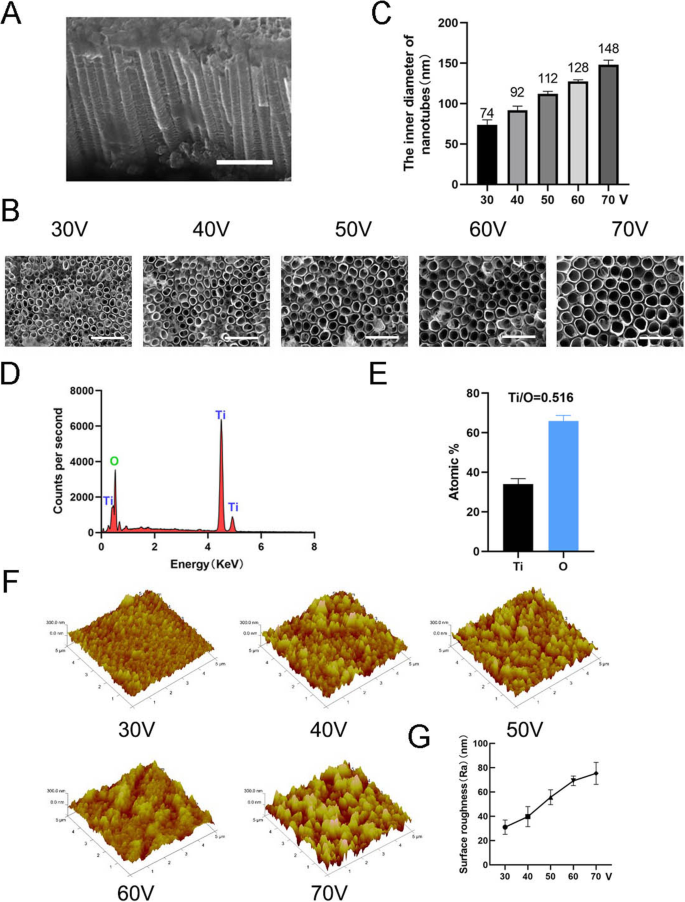
Caracterización superficial de nanotubos. un Vista lateral de nanotubos. Barras de escala:1 μm. b Vista superior de nanotubos. Barras de escala:500 nm. c El diámetro interior de los nanotubos a cinco voltajes constantes diferentes (30, 40, 50, 60, 70 V). d La composición del elemento químico EDS de las estructuras de nanotubos. e La relación de elementos de Ti y O. f Imágenes de microscopía de fuerza atómica (AFM) de estructuras de nanotubos. g La rugosidad superficial media (Ra) de los nanotubos
TiO 2 Diferenciación osteogénica de las CMM inducida por nanotubos
Después de 7 días de osteoinducción, la tinción de ALP se llevó a cabo por primera vez para evaluar la diferenciación osteogénica de las CMM. Los resultados de la tinción mostraron que las MSC cultivadas en TiO 2 Los nanotubos tenían una actividad ALP más alta que las células cultivadas sobre un sustrato de titanio liso (grupo de control) (Fig. 3a). El análisis estadístico del área de tinción demostró que la capacidad de los nanotubos para inducir la diferenciación osteogénica se incrementó significativamente en comparación con el grupo de control. Mientras tanto, observamos una tendencia de que dentro del rango de diámetros de este experimento, cuanto mayor es el diámetro del TiO 2 nanotubos, mayor es la capacidad de inducir la diferenciación osteogénica (Fig. 3b). Por lo tanto, el grupo de 70 V se utilizó en experimentos posteriores para mostrar mejor los resultados. A continuación, analizamos la expresión del gen osteogénico en los días 3 y 7. MSC cultivadas en TiO 2 Los nanotubos durante 3 y 7 días mostraron ambos una promoción significativa de la expresión de genes osteogénicos (RUNX2, ALP, OCN y OSX) en comparación con el grupo de control (Fig. 3d-g). Los resultados de la transferencia Western confirmaron que la expresión de proteínas de RUNX2 y OSX también aumentó después de 7 días de osteoinducción (Fig. 3c). Curiosamente, encontramos que la F-actina estaba regulada al alza en el TiO 2 grupo de nanotubos. Por lo tanto, era obvio que TiO 2 Los nanotubos dirigieron las MSC hacia la diferenciación de osteoblastos, que estaba relacionada con el diámetro de los nanotubos. Nuestros resultados también sugirieron la participación de F-actina en este proceso.
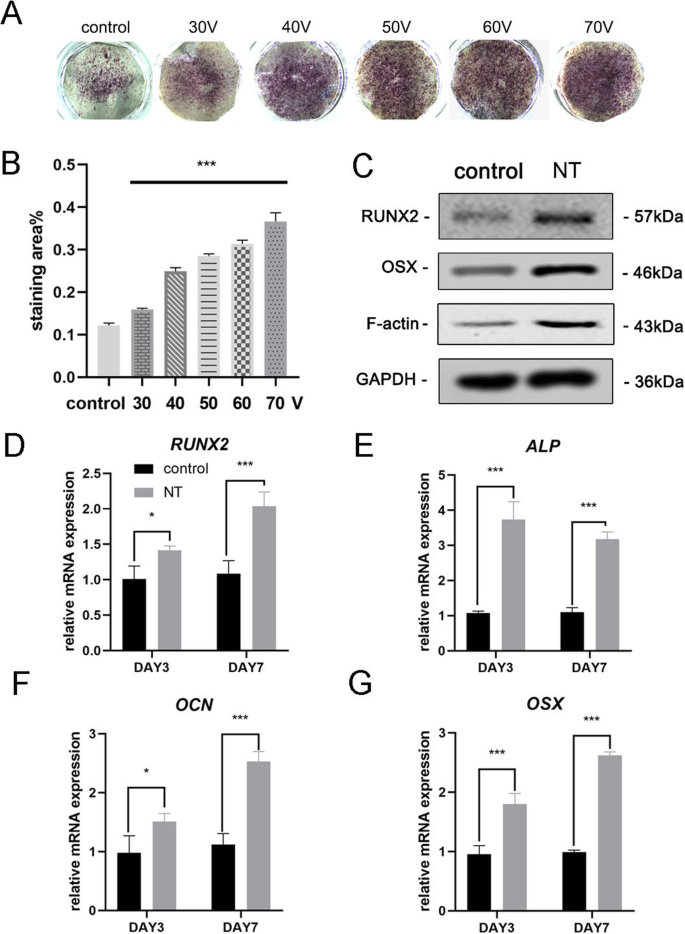
TiO 2 Los nanotubos mejoraron la expresión génica osteogénica de las BMSC. un Tinción ALP del sustrato liso de titanio y cinco sustratos de nanotubos diferentes. Las células se indujeron con medio osteogénico durante 7 días. b El análisis estadístico del área de tinción se realizó usando ImageJ. c Las proteínas asociadas a la osteogénesis (RUNX2 y OSX) y la actina F en las MSC se analizaron mediante transferencia Western el día 7. La expresión de ARNm de RUNX2 ( d ), ALP ( e ), OCN ( f ) y OSX ( g ) en los días 3 y 7, analizados por qRT-PCR. NT el grupo de nanotubos. Los datos representan la media ± DE de tres muestras. * P <0.05, ** P <0.01 y *** P <0,001
Diferenciación de osteoblastos mediada por F-actina de MSC en TiO 2 Nanotubos
Para explorar más a fondo si la F-actina estaba involucrada en la diferenciación celular inducida por la nanotopografía, utilizamos dos reactivos, jasplakinolide (Jasp) y citocalasina D (Cyto D), para regular la polimerización de F-actina de forma positiva y negativa, respectivamente. Las microfotografías confocales de la tinción con rodamina-faloidina mostraron que la F-actina en el grupo de tratamiento con Cyto D estaba casi despolimerizada y que raras veces se veían estructuras fibrosas, mientras que Jasp estabilizó y polimerizó la F-actina, verificada por estructuras en forma de haz más distintas y más brillantes que las observadas. en el grupo de control (Fig. 5a). Además, el análisis de transferencia Western también confirmó que la expresión de la proteína de F-actina se vio afectada, lo que demostró que tanto Cyto D como Jasp actuaron como se esperaba (Fig. 5b). El ensayo de proliferación celular mostró que Cyto D inhibía significativamente la proliferación celular, mientras que Jasp promovía el crecimiento celular (Fig. 4a). Los resultados del recuento de células fueron consistentes con este hallazgo (Fig. 4b).
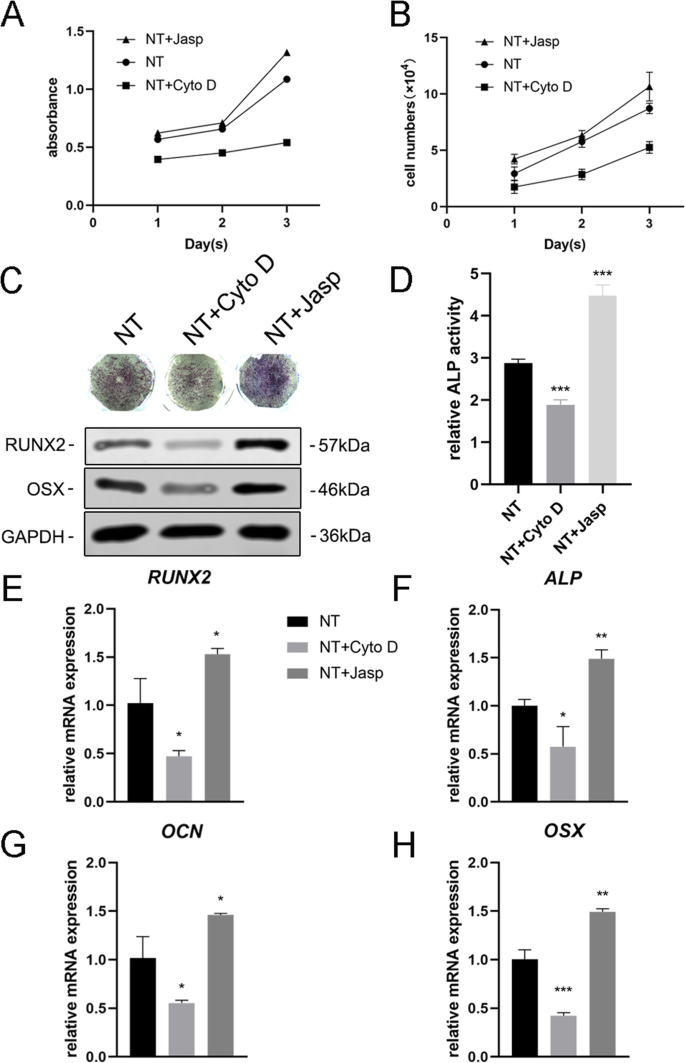
El ensamblaje de F-actina regulaba la expresión de genes osteogénicos en BMSC. un , b La proliferación celular después del tratamiento con Cyto D y Jasp se determinó mediante el ensayo CCK-8 o un contador celular automático en los días 1, 2 y 3. c , d Se realizó una tinción de ALP y un análisis de la actividad de ALP para evaluar la expresión de ALP en las MSC después del tratamiento farmacológico durante 3 días. El área de tinción se analizó usando ImageJ. c , e - h Se utilizaron Western blot y qRT-PCR para comparar los cambios de los marcadores relacionados con la osteogénesis en el grupo NT + Cyto D y el grupo NT + Jasp con los del grupo de control (sin tratamiento farmacológico). NT el grupo de los nanotubos. Los datos representan la media ± DE de tres muestras. * P <0.05, ** P <0.01 y *** P <0,001
A continuación, evaluamos la capacidad de las MSC para diferenciarse en osteoblastos para investigar si la F-actina mediaba este proceso. Primero detectamos ALP como un marcador temprano de osteogénesis. En comparación con el grupo de control, el tratamiento con Cyto D redujo la expresión de ALP y su actividad, mientras que en el grupo de tratamiento con Jasp se incrementó (Fig. 4c, d). De acuerdo con este resultado, el tratamiento con Jasp dio como resultado un aumento en los niveles de proteína de RUNX2 y OSX, mientras que Cyto D tuvo el efecto opuesto (Fig. 4c). De acuerdo con esto, los niveles de expresión de ARNm de genes osteoespecíficos, incluidos RUNX2, ALP, OCN y OSX, mostraron la misma tendencia después del tratamiento farmacológico (fig. 4e-h). Sobre todo, estos datos indicaron que la F-actina desempeñaba un papel importante en el proceso de diferenciación osteogénica de las CMM inducidas por TiO 2 nanotubos. La promoción de la despolimerización de F-actina inhibió la diferenciación de osteoblastos inducida por nanotopografía, mientras que la estabilización y polimerización de F-actina mejoró la diferenciación de osteoblastos.
Diferenciación de osteoblastos regulada por actina F de MSC en TiO 2 Nanotubos a través de MKL1 y YAP / TAZ
Para diseccionar el mecanismo subyacente que involucra a F-actina en la regulación del destino de las MSC, investigamos proteínas / moléculas que interactúan directamente con F-actina o afectan la polimerización de F-actina. Primero, intentamos identificar cómo la nanotopografía afectaba el equilibrio entre F-actina y G-actina. TiO 2 Los nanotubos como señal física difieren de las señales químicas permeables a la membrana y, por lo tanto, deben utilizar algunos componentes de la membrana celular para transmitir estímulos a las células. La evidencia acumulada indica que el complejo de adhesión focal, que incluye integrina, talina, quinasa de adhesión focal (FAK), vinculina (VCL), tensina y otras proteínas, funciona como un portador de señales, que informa a las células sobre el estado de la matriz extracelular (ECM). ) y por tanto afecta a su comportamiento biológico [22, 23]. Más importante aún, la F-actina se une a las integrinas a través de tales complejos de adhesión focal y, por lo tanto, forma enlaces mecánicos entre los haces de actina intracelulares y la MEC [24]. En consecuencia, analizamos a continuación la expresión de los componentes del complejo de adhesión focal. Los resultados mostraron que la expresión de proteínas y ARNm de VCL y FAK eran consistentes con el cambio de F-actina, lo que indica que el complejo de adhesión focal estaba involucrado en el proceso de diferenciación osteogénica de MSC inducida por TiO 2 nanotubos (Figs. 5b y 6a, b). Además, también encontramos que RhoA, una pequeña proteína GTPasa en la familia Rho de GTPasas, estaba regulada al alza en el grupo de tratamiento con Jasp e inhibida por Cyto D (Figuras 5b y 6a, b). RhoA es una importante molécula de transducción de señales en sentido ascendente en la vía MAPK, y podría ser regulada por FAK [25, 26]. La función principal de RhoA es promover la polimerización y estabilidad de las fibras de tensión (F-actina) y el ensamblaje del complejo de adhesión focal [27]. En conjunto, estos datos sugirieron que TiO 2 Los nanotubos podrían influir en la polimerización de F-actina a través del complejo de adhesión focal y RhoA.
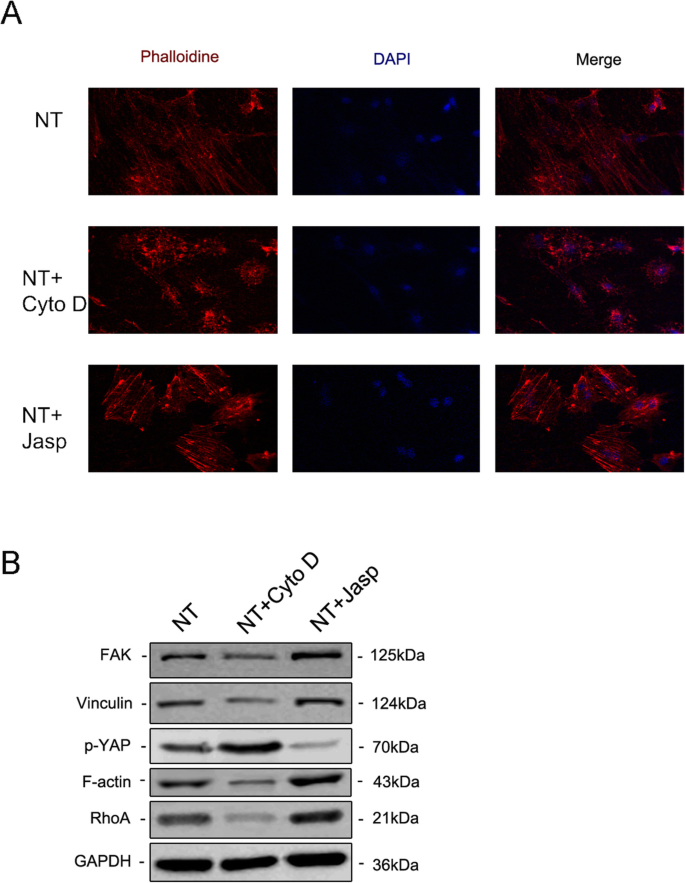
La tinción por inmunofluorescencia reveló el nivel de F-actina mediante la tinción con faloidina conjugada con rodamina ( a ). La expresión de proteínas de FAK y VCL contenidas en el complejo de adhesión focal, RhoA y YAP fosforilado se investigaron mediante transferencia Western ( b ). NT el grupo de nanotubos

Effect of F-actin assembly on FAK (a ), vinculin (VCL) (b ), RhoA (c ), MKL1 (d ), YAP (e ), and TAZ (f ) gene expression in MSCs. NT the nanotubes group. Data represent the mean ± SD of three samples. * P <0.05, **P <0.01, and ***P <0.001
But how does F-actin regulate cell fate? Most studies have demonstrated that F-actin is involved in cell migration, cell division, endocytosis, and especially tumor cell invasion [28,29,30]. Few studies have suggested that F-actin could also regulate cell differentiation, let alone its specific molecular mechanism [31, 32]. Consequently, we searched for articles that mentioned the F-actin changes and found that YAP/TAZ, two closely related transcriptional co-activators in the Hippo signaling pathway, which shuttle between the cytoplasm and the nucleus, may serve as mechanotransducers in regulating MSC differentiation [33,34,35]. In addition, we also found that MKL1, a key regulator of smooth muscle cell differentiation, which interacts with the transcription factor serum response factor, could bind to G-actin and also circulate between the cytoplasm and the nucleus [21, 36]. Our results ultimately identified the involvement of YAP/TAZ and MKL1 in nanotube-induced osteoblast differentiation mediated by F-actin (Figs. 5b and 6d–f). Interestingly, the protein expression of phosphorylated YAP showed the opposite trend, indicating that not only was the expression of YAP changed, but the phosphorylation of YAP was also changed by Cyto D and Jasp (Fig. 5b). This result was consistent with the report that the phosphorylation of YAP/TAZ could be sequestrated in the cytoplasm [35].
In summary, our results preliminarily demonstrated that F-actin regulated osteoblast differentiation of MSCs on TiO2 nanotubes through MKL1 and YAP/TAZ (Fig. 7).

Schematic representation of F-actin assembly induced by nanotubes, and the putative role of MKL1 and YAP/TAZ in acting as the downstream mediators of F-actin signaling to regulate gene expression
Discusión
Titanium and titanium alloys are the most widely used metal materials in orthopedic clinical implants due to the good properties of titanium [3]. However, aseptic loosening is still an urgent problem to be solved and improved, and the key is likely to lie in improving the integration of the implant and the host bone. Previous studies have shown that surface coating and modification or immobilization of biofunctional molecules will be beneficial to osseointegration [37]. Recently, the surface topography of implants has attracted the attention of many researchers thanks to studies into the cell response to physical cues [9,10,11, 13, 38]. In this study, we demonstrated the ability of nanotubes to promote osteogenic differentiation of MSCs, and this ability was enhanced with increasing inner diameter of the nanotubes (30–70 V). This will help guide the diameter of the nanotubes on the surface of the implants.
As a topographical structure, nanotubes first change the physical properties of the material, such as adsorption capacity and electrical and thermal conductivity. These physical properties determine their application in the industrial field. For example, most high-voltage power transformers need to be filled with insulating material, which is usually transformer oil or insulating gas. When the insulation of a transformer fails due to overheating and partial discharge, a serious discharge accident will occur. Therefore, finding an effective method that accurately detects the concentration and types of dissolved gases or insulating gas decomposition components in a transformer is necessary to monitor the operating state of the transformer [8, 39,40,41]. The traditional approach is to look for materials with good gas adsorption in transition elements, which are rich in d electrons, such as Pd(1 1 1) [39]. Nowadays, nanotubes are widely studied for their good gas adsorption properties. He et al. found that CuO-BNNT was suitable for the adsorption of C2 H 2 , because of its stronger adsorption on C2 H 2 [8]. Meanwhile, TiO2 itself can be a gas-sensing material. Gui et al. found that Co-doped TiO2 further enhanced gas adsorption capacity and exhibited a superior adsorption ability and conductivity change toward C2 H 4 molecules [40]. Consistent with this study, Mn-doped graphene also exhibited enhanced conductivity and superior capability of C2 H 2 and CO detection than pristine graphene [41]. The above research indicates that the TiO2 nanotubes prepared in our experiment have a potential application in the field of monitoring the operative state of a transformer. However, the adsorption capacity and electrical conductivity of the nanotubes to gases need to be further studied, especially whether these properties are enhanced after doping with transition elements (e.g., Mn).
In addition to changing the physical properties of a surface, nanoscale morphology also affects the biological behavior of the cells attached to it. Cells first adhere to the surface of the material and then migrate, proliferate, and differentiate. Compared to a flat surface, the hollow structure of the nanotubes provides fewer adhesion sites for cells. Therefore, in order for the cells to adhere to the nanotube surface steadily and maintain the biomechanical balance within the cell, the focal adhesion complex begins to assemble and mature, and F-actin becomes strong and stable.
F-actin, a linear polymer microfilament consisting of G-actin monomers, is one of the three major components of the cytoskeleton. As a mechanical-loading structure, F-actin is generally believed to be involved in cell division, cell migration, endocytosis, and tumor cell invasion [28,29,30], but some recent studies showed that it can also affect cell differentiation [31,32,33, 36]. For example, actin cytoskeletal depolymerization by simvastatin induces chondrocyte differentiation [31], and actin depolymerization enhances adipogenic differentiation in human stromal stem cells [32]. Our results also revealed that, compared with the control group, MSCs cultured on nanotubes had higher F-actin levels and a more obvious fibrous structure. Meanwhile, promotion of F-actin polymerization by Jasp enhanced osteogenic differentiation, while the depolymerization of F-actin inhibited osteogenic differentiation, suggesting that F-actin mediates TiO2 nanotube-induced osteoblastic differentiation of MSCs.
F-actin can be regulated by Rho GTPases, members of the Ras superfamily [23, 42], and Rho can induce actin reorganization through at least two effectors, ROCK and Dia. ROCK is activated by binding to Rho-GTP and then myosin light chain (MLC), the substrate of ROCK, plays an important role in F-actin assembly. ROCK inhibits the activity of MLC phosphatase, leading to an increase in MLC phosphorylation, which stimulates the ATPase activity of myosin II and promotes the assembly of F-actin. In addition, ROCK also targets LIM kinase (LIMK). Phosphorylated LIMK inactivates cofilin by phosphorylation, which can disassemble F-actin in its active state. Another effector is Dia, a member of the formin-homology (FH) family of proteins which contains two FH domains. These domains contain multiple proline-rich motifs which bind to the G-actin-binding protein, profilin. This interaction contributes to actin polymerization and F-actin organization [42]. We detected one of the Rho GTPases, RhoA, and found that the expression of RhoA was consistent with the level of F-actin. However, we were unable to clearly describe how the nanotubes regulate the expression of RhoA, because there are many other regulators, including integrin signaling, other adhesion receptors, G protein-coupled receptors (GPCRs), soluble factors such as LPA, receptor tyrosine kinase signaling, and so on [43].
Knowing that F-actin can be regulated by RhoA, we next asked what role focal adhesion played in this process, because focal adhesion complexes, containing integrins, talin, vinculin, paxillin, and focal adhesion kinase (FAK), are formed and mature when cells attach to the surface of nanotubes. Integrins are transmembrane heterodimers that couple the ECM to the other focal adhesion proteins so as to facilitate cell attachment. They not only act simply as hooks but also transmit to the cell critical signals about the nature of its surroundings, which along with other signals such as EGFR, prompt the cell to make decisions about its biological behaviors. These signals are further transmitted to F-actin, which is directly connected to the focal adhesion complexes. On the one hand, the nanoscale morphology causes focal adhesion complex assembly and maturation. On the other hand, kinases such as FAK and Src kinase family members will recruit molecules such as CRK to self-regulate the assembly and maturation of focal adhesion complexes [44,45,46]. Our results demonstrated that the formation and maturation of focal adhesion complexes were impaired by F-actin depolymerization, suggesting that there was a feedback from focal adhesion complexes to actin assembly in line with published reports.
However, it should not be ignored that these proteins contained in focal adhesion complexes have the function of signal transduction [47]. That is to say, nanotubes may directly regulate gene expression through signal cascades, and F-actin may just participate in or be affected by this process. For instance, the dual kinase complex of FAK and Src can regulate Rho GTPases such as RhoA. This shows that nanotubes can regulate RhoA through integrins and the FAK/Src complex. In addition Src, a non-receptor tyrosine kinase protein, can activate Ras (small GTPase) by phosphorylating FAK at tyrosine residue 925 [47, 48]. Then, Ras activates numerous biochemical pathways, including the well-studied MAPK pathway and the PI3K/AKT/mTOR pathway. In the MAPK pathway, Ras activates c-Raf, followed by mitogen-activated protein kinase kinase (MAP2K) and then MAPK1/2, also known as extracellular signal-regulated kinase (ERK). ERK in turn activates transcription factors such as serum response factor (SRF) and c-Myc that are involved in regulating growth and differentiation [49]. What is more, Runx2, a key transcription factor in osteogenic differentiation, can also be regulated by ERK [50], and our previous study confirmed that mechanical strain promoted osteogenic differentiation of BMSCs through the FAK-Erk1/2-Runx2 pathway [17]. Therefore, we cannot rule out that ERK plays a role in nanotube-induced osteogenic differentiation and further study is still needed.
So what exactly is the role of F-actin in inducing differentiation of nanotubes, because its change can affect cell differentiation? One possibility is that the change of F-actin assembly can inversely regulate the level of FAK so as to induce osteogenic differentiation through the FAK-Erk1/2-Runx2 pathway as described above, because in our results, focal adhesion complexes and actin polymerization showed the same trends of change, indicating that they act as a whole in response to the extracellular environment. However, some other possibilities also exist, and a number of articles have shown that MKL1 and YAP/TAZ act downstream of the actin dynamic balance [20, 51,52,53,54]. Both of them shuttle between the cytoplasm and the nucleus, and may help to transduce signals from the cytoskeleton to the nucleus.
MKL1, also termed myocardin-related transcription factor A, is sensitive to changes in G-actin levels. When cytoplasmic G-actin levels increase, monomeric G-actin binds to MKL1 and prevents it from binding to SRF and activating transcription. SRF target genes include actins such as smooth muscle actin (SMA) as well as other actin-binding proteins, including immediate early genes like c-fos and egr1. Recent studies have demonstrated that changing SRF activity could regulate adipogenesis by activating the adipogenesis transcription factor peroxisome proliferator-activated receptor γ (PPARγ), and also regulate bone formation via IGF-1 and Runx2 signaling [55, 56].
YAP and TAZ are two transcriptional coactivators in the Hippo signaling pathway, identified as an important regulatory pathway that restricts cell proliferation, thereby controlling organ size and morphogenesis [20]. Large tumor suppressor genes 1 and 2 (LATS1/2) phosphorylate them, thereby creating a binding site for 14-3-3 proteins, the binding of which prevents their nuclear import [53, 54]. As a consequence, phosphorylated forms of YAP/TAZ are sequestered in the cytoplasm, preventing the expression of genes like Ctgf and Areg. In addition, some studies have shown that YAP/TAZ can interact with T-box 5 (TBX5), RUNX2, and p73 to regulate gene expression [57,58,59]. Further, cell adhesion to cell matrix proteins has been shown to trigger YAP nuclear localization through an integrin/FAK/Src axis. In our study, the results suggested that this pathway was possibly involved in nanotube-induced differentiation. Further study into the downstream mediators of the integrin/FAK/Src axis should be carried out to clarify the specific mechanism.
On the other hand, more and more studies illustrate that F-actin interacts with Hippo signaling, and somehow inhibits the phosphorylation of YAP [54, 60], which is consistent with our experimental results that promoting F-actin polymerization reduces the expression of phosphorylated YAP. We hypothesize that ATP involved in the process of the transformation between G-actin and F-actin may also play an important role in the phosphorylation of YAP, which is yet to be studied.
After understanding the above possible molecular mechanisms, we can try to explain some of the experimental phenomena found in this study. Our results revealed that the larger the diameter of the nanotubes, the stronger the ability of the nanotubes to promote osteogenic differentiation. This is consistent with previous research [61, 62]. The reason for this phenomenon is that the larger the diameter of the nanotubes, the less adhesion sites they can provide to the cells, and the greater the assembly and maturity of focal adhesion complexes. Along with these, stress fibers made of F-actin will have greater strength and stability. These structures enhance the signaling that promotes osteogenic differentiation. Predictably, however, this effect is significantly reduced when the nanotubes become too large in diameter, making it difficult for the cells to adhere to the surface [12]. Similarly, when the height of the nanotubes is inconsistent, the differences in height can result in a change of adhesion site and rearrangement of the cytoskeleton, which will further affect cell differentiation. Intriguingly, even flat surface materials without nanotube modification can induce changes in cell differentiation. A number of studies have demonstrated that focal adhesion formation and stress fiber organization are regulated by substrate stiffness [63,64,65], and YAP/TAZ also plays an important role in this process. Therefore, it is obvious that the integrins–FAs (focal adhesions)–F-actin axis plays a role in the transduction of physical signals into intracellular chemical signals.
In summary, our results demonstrated that F-actin regulates osteoblastic differentiation of mesenchymal stem cells on TiO2 nanotubes through MKL1 and YAP/TAZ, whose target genes partly explained the proliferation and differentiation of MSCs. We know that there is no single change in the signal network and any change is regulated by numerous molecules and proteins. One type of biological behavior must be the result of the regulation of a series of signaling pathways. Nanotubes induce cell differentiation by triggering a complex network of signals, including integrins, proteins contained in focal adhesion complexes, FAK, Src, Rho GTPase, the MAPK pathway, the Hippo pathway, and other reported signaling pathways. At least as important, there are many signal cycles in the signal network and a downstream signal can regulate the upstream signal via feedback. In this study, we found that vinculin and FAK can be regulated backwards by F-actin assembly, increasing the uncertainty of molecular function. Therefore, more details of the molecular mechanism await further study.
Conclusiones
Nuestros resultados mostraron que TiO 2 nanotubes promoted the osteogenic differentiation of MSCs, and this ability was enhanced with the increasing diameter of nanotubes within a certain range (30–70 V). F-actin mediated nanotube-induced cell differentiation through MKL1 and YAP/TAZ, providing a novel insight into the study of cell differentiation.
Disponibilidad de datos y materiales
Los conjuntos de datos utilizados y analizados durante el estudio actual están disponibles del autor correspondiente a solicitud razonable.
Abreviaturas
- MSCs:
-
Mesenchymal stem cells
- SEM:
-
Microscopía electrónica de barrido
- EDS:
-
X-ray energy dispersive analysis
- AFM:
-
Microscopía de fuerza atómica
- ALP:
-
Fosfatasa alcalina
- Cyto D:
-
Cytochalasin D
- Jasp:
-
Jasplakinolide
- VCL:
-
Vinculin
- FAK:
-
Focal adhesion kinase
- BCA:
-
Bicinchoninic acid
- PVDF:
-
Polyvinylidenedifluoride
- Runx2:
-
Runt-related transcription factor 2
- Osx:
-
Osterix
- OCN:
-
Osteocalcin
- YAP:
-
Yes-associated protein
- MKL1:
-
Megakaryoblastic leukemia 1
- FBS:
-
Suero fetal bovino
- ECM:
-
Extracellular matrix
- MLC:
-
Myosin light chain
- LIMK:
-
LIM kinase
- FH:
-
Formin-homology
- GPCR:
-
G protein-coupled receptors
- MAP2K:
-
Mitogen-activated protein kinase kinase
- ERK:
-
Extracellular signal-regulated kinase
- SRF:
-
Serum response factor
- SMA:
-
Smooth muscle actin
- PPARγ:
-
Peroxisome proliferator-activated receptor γ
- LATS1/2:
-
Large tumor suppressor gene 1 and 2
- TBX5:
-
T-box 5
Nanomateriales
- DIÓXIDO DE TITANIO - TiO2 - Precio, mercado y análisis
- Hacia los nanofluidos de TiO2:Parte 1:Preparación y propiedades
- Las nanotecnologías desactivan las células madre cancerosas
- Hacia los nanofluidos de TiO2:Parte 2:Aplicaciones y desafíos
- Efecto de la distribución de nanopartículas de oro en TiO2 sobre las características ópticas y eléctricas de las células solares sensibilizadas por colorante
- Síntesis de nanocristales de ZnO y aplicación en células solares de polímero invertido
- El precursor de titanio óptimo para la fabricación de una capa compacta de TiO2 para células solares de perovskita
- Influencia de la rigidez elástica y la adherencia de la superficie en el rebote de nanopartículas
- Influencia de nanopartículas de agricultura con diferentes tamaños y concentraciones incrustadas en una capa compacta de TiO2 en la eficiencia de conversión de las células solares de perovskita
- Un nuevo material de conversión ascendente de Ho3 + -Yb3 + -Mg2 + TiO2 tri-dopado y sus aplicaciones a las células solares de perovskita
- Influencia de los nanotubos de carbono y sus derivados en las células tumorales in vitro y parámetros bioquímicos, composición de la sangre celular in vivo