


Las nanotecnologías desactivan las células madre cancerosas
Resumen
Una de las tareas de la oncología actual es la identificación de las células madre cancerosas y la búsqueda de medios terapéuticos capaces de inhibirlas de forma específica. El artículo presenta los datos sobre las características fenotípicas de las células del carcinoma de Ehrlich como modelo de crecimiento tumoral conveniente y fácil de seguir. La evidencia de células madre cancerosas como parte del carcinoma de Ehrlich y la importancia de CD44 + y CD44 - Se demostraron subpoblaciones en el mantenimiento del crecimiento de este tipo de tumor. Una alta (diez veces) actividad tumorigénica del carcinoma de Ehrlich CD44 + células en comparación con CD44 - las células fueron probadas. En este par de comparación, el CD44 + las células tenían un mayor potencial de generar en la cavidad peritoneal de CD44 alto , CD44 + CD24 - , CD44 + CD24 + subpoblaciones de células, destacando la presencia de células madre cancerosas en un grupo de CD44 + células.
En este estudio, la capacidad de los nanocomplejos híbridos sintetizados, que comprenden las nanopartículas de ortovanadatos de tierras raras GdYVO 4 :Eu 3+ y se estableció colesterol para inhibir el crecimiento tumoral y aumentar la supervivencia de los animales con tumores. Cada uno de sus componentes hace una contribución especial al efecto inhibidor de tumores. El tratamiento de las células del carcinoma de Ehrlich con un complejo híbrido de dos componentes dio como resultado una reducción máxima en la concentración de CD44 más tumorigénico alto células con aumento simultáneo en el número de CD117 + células que disminuyeron la intensidad del crecimiento tumoral en un 74,70 ± 4,38% en comparación con el control.
Antecedentes
El problema del crecimiento maligno sigue siendo uno de los más urgentes en medicina. En las últimas décadas, ha habido algunos avances en el desarrollo de nuevos tratamientos para el cáncer. Esto se debe a la revisión del concepto clásico de cáncer y al descubrimiento de las células madre cancerosas (CSC), que son capaces de autorrenovarse ilimitadamente y pueden identificarse mediante una serie de marcadores fenotípicos. La mayoría de estas células son resistentes a las radio y quimioterapias, lo que provoca la recaída del crecimiento maligno y la metástasis. Hay nuevos métodos dominados de terapia contra el cáncer, a saber, que permiten inactivar selectivamente las células tumorales con un daño mínimo al tejido normal [1].
Las CSC fueron identificadas y descritas por primera vez en 1997 por el equipo de M. Dick [2]. Los autores investigaron una leucemia mieloide aguda en la que la subpoblación de 0,01% a 1% de la población total de células podría causar leucemia cuando se trasplantan a ratones NOD / SCID (inmunodeficiencia combinada grave diabética no obesos) inmunodeficientes. Estas células inductoras de tumores se caracterizaron fenotípicamente como CD34 + CD38 - . En 2003, M. Al-Hajj y M.S. Wicha logró identificar las CSC en una forma sólida de cáncer de mama humano (BC) [3]. Se ha encontrado que la población no separada de cáncer de mama primario exhibió un potencial tumorigénico en el 100% de los casos (10/10) cuando se administró a ratones NOD / SCID a una concentración de 5 × 10 4 células / ratón. Reducir la concentración de células administradas a 1 × 10 4 células / ratón disminuyeron su actividad tumorigénica 4 veces (3/12) [3]. CD24 + CD44 + fracción cuando se administra en varias dosis (2 × 10 4 hasta 100 células / ratón) no permitió el crecimiento del tumor. Adjunto, CD44 + CD24 - / baja subpoblación poseía una actividad tumorigénica significativamente mayor, demostrando la formación de tumores en el 100% de los casos cuando se administraba 10 3 células / ratón. La capacidad más pronunciada para formar tumores fue inherente a la subpoblación de CD44 + CD24 - / lo ESA + fenotipo. La administración a ratones de solo 200 de estas células resultó en la formación de tumores sólidos en el 100% (4/4) 5 meses después de su inyección [3]. Estos estudios fueron continuados por Ponti D. et al., Quienes demostraron la capacidad de ciertas poblaciones de muestras de biopsias de cáncer de mama para formar las mamasferas in vitro en cultivos libres de suero [4]. La mayoría de las células de las mamasferas obtenidas eran de CD44 + / CD24 - / bajo fenotipo así como un potencial tumorigénico aumentado in vivo cuando se administra a ratones SCID (inmunodeficiencia combinada grave). La capacidad de formar tumores en esta subpoblación fue 1000 veces mayor que la de la línea tradicionalmente trasplantada de carcinoma de mama MCF7 [4]. Sin embargo, los autores han demostrado que solo el 20% de CD44 + CD24 - / bajo las células tenían la capacidad de autorrenovarse. Esto puede deberse a la heterogeneidad de esta subpoblación, a saber, la presencia de marcadores adicionales (ESA, ALDH), que determinan la función de las células y también puede estar relacionado con la tasa de expresión de CD44. Los trabajos publicados durante los últimos años muestran que las CSC con una alta expresión del marcador (CD44 alta ) tienen la mayor actividad tumorigénica [5, 6]. En implantación ortotópica de 5 × 10 5 CD44 alto -RAS-transformado y CD44 bajo células a ratones NOD / SCID, se ha encontrado que una subpoblación CD44 baja poseía baja tumorigenicidad (el tumor se formó en el 30% de los casos), mientras que el CD44 alto las células eran capaces de formar tumores en el 100% de los casos [6].
Resumiendo los datos publicados, la fila de diferenciación de subpoblaciones de células de cáncer de mama se puede representar de la siguiente manera:
$$ \ mathrm {C} \ mathrm {D} {44} ^ {\ mathrm {alto}} \ to \ mathrm {C} \ mathrm {D} {44} ^ {+} \ mathrm {C} \ mathrm { D} {24} ^ {\ hbox {-}} \ to \ kern0.5em \ mathrm {C} \ mathrm {D} {44} ^ {+} \ mathrm {C} \ mathrm {D} {24} ^ {+} \ to \ mathrm {C} \ mathrm {D} {44} ^ {\ hbox {-}} \ mathrm {C} \ mathrm {D} {24} ^ {+} $$Varias celdas que llevan otros marcadores, en particular Sca-1 + reclama la etapa de CSC. Los datos sobre una reducción del crecimiento tumoral en los ratones knock-out para Sca-1 indican a favor de la hipótesis del papel iniciador del tumor de Sca-1 + células en una etapa temprana de tumorigénesis [7]. Recientemente, mucha más atención de los investigadores ha sido atraída no solo por las CSC, sino también por las células que forman su microambiente regulador accesorio. El CD117 + las células, que tradicionalmente se detectan en un grupo de células madre sanguíneas, merecen una atención particular entre ellas [8]. La población total de células de carcinoma de mama humano comprende los denominados fibroblastos del estroma asociados al carcinoma con CD117 + fenotipo. Apoyan el crecimiento del tumor, promoviendo su angiogénesis [9, 10]. Una suposición de la presencia en la población de células madre de carcinoma de Ehrlich (CE), estudio del potencial tumorigénico de CD44 + fracción y función de CD117 + células para mantener el desarrollo del tumor requiere una evidencia adicional.
La mayoría de los experimentos para pasar CSC in vivo se realizaron en ratones SCID o NOD / SCID. Estos ratones no responden con una reacción inmune al xenotrasplante de células humanas. Está en marcha la búsqueda de modelos experimentales adecuados y relevantes para estudiar y evaluar la actividad antitumoral de diversos agentes terapéuticos. Uno de ellos es la línea de células tumorales trasplantadas in vivo de EC, que se obtuvo de un cáncer de mama espontáneo de ratones [11]. Sin embargo, prácticamente no existen publicaciones en cuanto a la composición de subpoblaciones de células CE y sus características fenotípicas, presencia de CSC y su importancia en el mantenimiento del crecimiento de este tipo de tumor. Teniendo en cuenta la similitud histogenética de EC y BC, se puede suponer que en el inicio y desarrollo del tumor simulado, pueden estar involucrados los mismos genes que controlan la proliferación de células cancerosas, así como vías bioquímicas similares que conducen a la expresión de proteínas marcadoras tumorales. Sin embargo, la suposición sobre la presencia de CSC en la población de CE y el estudio de su potencial tumorigénico necesita evidencia adicional, que fue uno de los objetivos del presente estudio.
Un problema no menos urgente de la oncología actual es encontrar los fármacos, no solo reconociendo específicamente sino también inactivando las CSC. El concepto mismo de comprensión del problema fue la base formada en la dirección del momento “teranóstica” (terapia + diagnóstico) [12]. En el marco de la teranóstica, se desarrollan enfoques tecnológicos de uso de medicamentos y herramientas de diagnóstico y terapia simultánea del cáncer. Una de las direcciones de la teranóstica es dirigir las nanopartículas de oro al sitio del tumor y después de la terapia fototérmica [13]. Otro enfoque para la identificación de células tumorales es el uso de puntos cuánticos, poderosos agentes de contraste óptico que permiten el seguimiento del tumor in vivo [14]. La capacidad de los puntos cuánticos para visualizar de forma no invasiva las células madre embrionarias humanas in vivo testifica a favor de su posible aplicación biomédica [15].
Recientemente se ha prestado mayor atención a los nanoluminóforos basados en materiales dieléctricos y semiconductores de zona amplia, activados con elementos de tierras raras, a saber, nanopartículas (NP) de metales de tierras raras (en particular, vanadio y sus compuestos) [16]. Estos materiales poseen una alta fotoestabilidad, un gran cambio de luminiscencia de Stokes, ausencia del efecto de centelleo y estabilidad de las características bandas de luminiscencia estrechas. Por lo tanto, se conocen los efectos antitumorales de los compuestos de vanadio. Entonces, se ha demostrado que un dicloruro de vanadio puede inhibir significativamente la proliferación celular como resultado de la acumulación en heterocromatina nuclear con la subsecuente inducción de aberración mitótica, supresión transitoria de mitosis, lo que lleva a la acumulación de células en S tardío y G 2 fases [17]. Prometedor para el tratamiento de tumores malignos puede ser el uso de nanocomplejos híbridos basados en NP de ortovanadatos de tierras raras GdYVO 4 :Eu 3+ y colesterol, desarrollado en el Instituto de Materiales de Centelleo de la Academia Nacional de Ciencias de Ucrania [18].
El propósito de su creación fue potenciar el efecto terapéutico de los agentes anticancerígenos debido a la presencia en la composición de nanocomplejos que tienen afinidad por las membranas de las células diana. Uno es el colesterol que se "retira" activamente del torrente sanguíneo mediante la proliferación de células cancerosas para construir las biomembranas. Esto se ve facilitado por la presencia en la superficie de un gran número de células tumorales SR-B1 (receptor eliminador, clase B tipo I) y receptores de caveolina-1 (Cav-1), que pueden unirse al colesterol sanguíneo libre [19]. .
Así, el objetivo de este trabajo fue identificar la composición de subpoblaciones de células CE, incluidas aquellas con signos de CSC, así como su actividad tumorigénica tras el pretratamiento con nanocomplejos híbridos.
Métodos
Los experimentos se realizaron en ratones Balb / C hembra de 8 meses. Los ratones se mantuvieron en condiciones estándar del vivero (temperatura ambiente de 20 ± 2 ° C, humedad relativa 50-70%, ciclo de luz-oscuridad 12:12 h). Todos los protocolos experimentales fueron aprobados por el Comité de Ética Animal del Instituto para Problemas de Criobiología y Criomedicina de la Academia Nacional de Ciencias de Ucrania, Kharkiv, Ucrania, (rec. N. ° 1 de 23.01.2017) y se ajustan a la Convención Europea sobre el Uso de Animales Experimentales (Estrasburgo, 1986), aprobado por el Primer Congreso Nacional de Ucrania en Bioética (Kiev 2004).
Cultivo de células EC in vivo
Se hicieron pases de células de carcinoma de Ehrlich (EC) en la cavidad peritoneal (PC) de ratones Balb / C. Las células EC criopreservadas en líquido ascítico se utilizaron como cultivo primario [20]. Después de la descongelación, las células CE se trasplantaron tres veces in vivo, para mitigar la influencia de los factores de congelación-descongelación y obtener por ellos las características morfológicas y funcionales de las células nativas [21]. "Estabilizado" por lo tanto, las células EC se inyectaron por vía intraperitoneal en una dosis de 3 × 10 6 células / ratón en 0,3 ml de solución salina y se cultivaron durante 7 días in vivo. Después de 7 días, los animales experimentales se retiraron del experimento bajo anestesia ligera con éter. Se extrajo líquido ascítico de PC con una jeringa a través de una aguja de 2,69 mm de diámetro interno y se colocó en un tubo de medición de 10 ml. El número absoluto de células se determinó magnificando el volumen del líquido ascítico acumulado en la cavidad peritoneal (ml) con el número de células EC contadas en la cámara de Goryaev. Un aumento en el número total hasta 35,00 × 10 7 Las células EC en PC de ratones hasta el día 7 fue un criterio de desarrollo de carcinoma [21]. En el futuro, las mismas células sirvieron como objeto de estudio.
Evaluación fenotípica de subpoblaciones de CE
Se realizó con un citómetro de flujo “FACS Calibur” (“Becton Dickinson,” EE. UU.) Utilizando anticuerpos monoclonales (EE. UU. “BD Biosciences”) contra CD44 (FITC) (no. 553133, clon IM7), CD117 (FITC) (no. 553354, clon 2B8) y Sca-1 (FITC) no. 553333, clon E13-161.7) y CD24 (PE) no. 553262, clon M1 / 69) de acuerdo con las instrucciones del fabricante. Como control, las muestras con la adición de anticuerpos monoclonales no inmunes marcados con FITC y PE de los mismos isotipos (“BD Biosciences”), no. 553988, clon A95-1 y no. 553989, clon A95-1), ya que se usaron anticuerpos contra el marcador probado. Se realizó una doble tinción inmunofenotípica utilizando anticuerpos monoclonales CD44 (FITC) y CD24 (PE). Las células con una fluorescencia media del marcador CD44 superior a 10 3 (según una escala logarítmica) fueron referidos a CD44 alto subpoblaciones. El registro y análisis de los resultados se realizaron con el software "WinMDi 2.9" (Joseph Trotter, La Jolla, EE. UU.).
Separación de CD44 + Fracción de células CE mediante clasificación inmunomagnética
Teniendo en cuenta que los CSC con un alto nivel de expresión del marcador CD44 (CD44 alto ), compuesta por una población heterogénea de CD44 + células, poseen la mayor actividad tumorigénica, se aislaron de una población de CE total con un clasificador magnético (BDTM Imagnet). Para aislar CD44 + Se utilizaron anticuerpos monoclonales primarios no marcados para el marcador CD44 (BD, 558739) y partículas magnéticas de IgG1 de ratón secundario-DM (BD, 557983) de acuerdo con el protocolo del fabricante. Pureza de separación para CD44 + células de la población total de CE fue del 90%.
Determinación de la actividad tumorogénica de las células de la población total y las de CD44 aislado + y CD44 - - Fracciones EC
Capacidad tumorigénica de la población total y CD44 aislado + y CD44 - Las fracciones de EC se analizaron comparativamente mediante el método de cultivo in vivo descrito anteriormente. La configuración experimental se muestra en la Fig. 1.
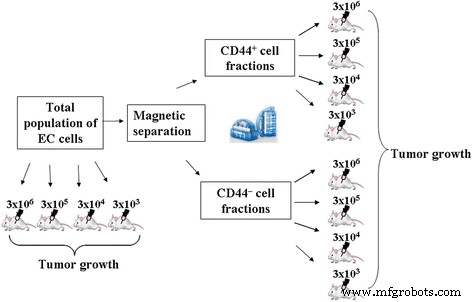
Diseño experimental al analizar comparativamente la capacidad tumorigénica de la población total y CD44 aislado + y CD44 - Fracciones EC
En el primer conjunto de experimentos, evaluamos la capacidad tumorigénica de la población total y el CD44 aislado + y CD44 - Fracciones de EC cuando se administran a animales en una dosis estándar utilizada para el inicio de EC (3 × 10 6 células en 0,3 ml de solución salina).
Los animales se dividieron en los siguientes grupos ( n =10):
-
Grupo 1.1:administración de la población total de células EC (3 × 10 6 células / animal)
-
Grupo 2.1:administración de CD44 + fracción de células EC (3 × 10 6 células / animal)
-
Grupo 3.1:administración de CD44 - fracción de células EC (3 × 10 6 células / animal)
En 7 días después de la inoculación en cada uno de los grupos experimentales, se contó un número total de células en PC de animales, se evaluaron las características fenotípicas de las células (como se describió anteriormente) y CD44 alto / CD117 + proporción de células EC determinada como la proporción de CD44 alta porcentaje a CD117 + células [22]. Potencial proliferativo de células de la población total de CE y CD44 aislado + y CD44 - fracciones se estimó basándose en los datos de la siguiente manera:factor de multiplicidad (MF) del excedente de población celular durante el tiempo de cultivo, M =N / N 0 ; y duplicación de tiempo (TD), TD =(log 2 2) * t / [log 2 ( N / N 0 )], donde t es el momento del cultivo celular (h), N es el número de celdas en t hora; N 0 es el número de celda inicial [23].
En el segundo conjunto de experimentos, hubo una dosis mínima estimada de las células administradas de la población total y CD44 aislado + y CD44 - Fracciones de EC, que inducen el crecimiento tumoral. Suspensión de células totales y CD44 aislado + y CD44 - Las fracciones de EC se administraron por vía intraperitoneal a ratones a dosis de 3 × 10 6 , 3 × 10 5 , 3 × 10 4 y 3 × 10 3 células por ratón en 0,3 ml de solución salina y se cultivaron durante 7 días en la PC.
Los animales utilizados en este conjunto de experimentos se han dividido en los siguientes grupos ( n =10):
-
Grupo 1.1:administración de la población total de células EC (3 × 10 6 celdasanimal)
-
Grupo 1.2:administración de la población total de células EC (3 × 10 5 células / animal)
-
Grupo 1.3:administración de la población total de células EC (3 × 10 4 células / animal)
-
Grupo 1.4:administración de la población total de células EC (3 × 10 3 células / animal)
-
Grupo 2.1:administración de CD44 + fracción de células EC (3 × 10 6 células / animal)
-
Grupo 2.2:administración de CD44 + fracción de células EC (3 × 10 5 células / animal)
-
Grupo 2.3:administración de CD44 + fracción de células EC (3 × 10 4 células / animal)
-
Grupo 2.4:administración de CD44 + fracción de células EC (3 × 10 3 células / animal)
-
Grupo 3.1:administración de CD44 - fracción de células EC (3 × 10 6 células / animal)
-
Grupo 3.2:administración de CD44 - fracción de células EC (3 × 10 5 células / animal)
-
Grupo 3.3:administración de CD44 - fracción de células EC (3 × 10 4 células / animal)
-
Grupo 3.4:administración de CD44 - fracción de células EC (3 × 10 3 células / animal)
En cada grupo experimental, se determinó el número total de células en CP y el de animales con desarrollo de ascitis 7 días después de la inoculación con CE.
Síntesis de nanocomplejos
Nanocomplejos híbridos que contienen nanopartículas esféricas (NP) (de 2 a 3 nm de diámetro) en una concentración de 1,30 g / ly colesterol de oveja en una concentración de 0,55 g / l (“Acros organics”, Bélgica) se sintetizaron en el Instituto de Materiales de Centelleo de la Academia Nacional de Ciencias de Ucrania (Járkov) como se informó [18]. NP basados en ortovanadatos de elementos de tierras raras GdYVO 4 :Eu 3+ de forma esférica en una concentración de 1,30 g / l se prepararon como se describe [24]. Las soluciones acuosas coloidales basadas en ortovanadatos han sido depuradas de impurezas mediante diálisis utilizando las membranas “Cellu Sep H1” de 3,5 KDa.
En el nanocomplejo híbrido, las NP cargadas negativamente se localizan a lo largo de la periferia de las partículas de colesterol debido a interacciones de van der Waals e hidrofóbicas. Los NP estabilizan los nanocomplejos mediante interacciones electrostáticas. Los tamaños de los nanocomplejos sintetizados no superan los 100 nm. Además, las NP exhiben propiedades antioxidantes y no están sujetas a oxidación. Este hecho contribuye al aumento de la resistencia de la dispersión acuosa de colesterol en relación con las especies reactivas de oxígeno. La estructura esquemática del nanocomplejo híbrido se muestra en la Fig. 2.
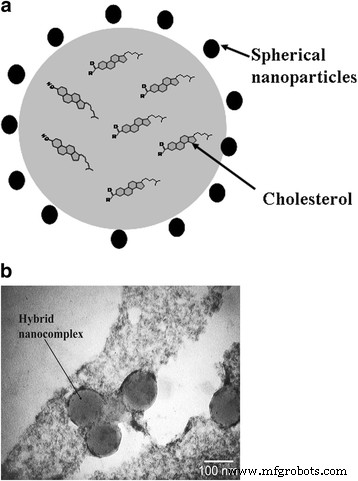
Nanocomplejo híbrido: a representación esquemática y b Microfotografía de microscopía electrónica de transmisión de nanocomplejos híbridos, obtenidos a partir de una solución acuosa de colesterol colocada en una red de carbono
Para registrar una acumulación de nanocomplejos híbridos en las células durante los estudios in vitro, se podría introducir adicionalmente el colorante fluorescente hidrófobo 1,1′-dioctadecil-3,3,3 ′, 3′-tetrametilindocarbocianina perclorato (DiI) en la dispersión acuosa de colesterol, permitiendo en espectroscopía luminiscente local evaluar la dinámica de integración del complejo en una membrana celular por la proporción de monómero - bandas de luminiscencia “J-agregado” [25]. Nuestros estudios anteriores han demostrado que el nanocomplejo híbrido no puede integrarse en más del 10% de las células de la población total de CE y prácticamente en todas las células de CD44 aislado + fracción que tiene el mayor potencial carcinogénico. Esto permite el uso de nanocomplejos híbridos en esta modificación (NP + colesterol + DiI) como método para identificar la acumulación local de nanocomplejos en las células cancerosas [26, 27].
Tratamiento previo de células CE con nanomateriales
La suspensión total de células CE con nanocomplejos híbridos o NP se incubó en una solución de glucosa al 5% (“Infusión” CJSC, Kyiv) a temperatura ambiente durante 3 h. Este tiempo de incubación se encontró anteriormente como óptimo para unir nanocomplejos a las células [26].
Se probaron las siguientes variantes de pretratamiento de células EC con nanomateriales:
-
Opción 1–900 μl de células EC (1 × 10 7 ) Se añadieron 100 μl de NP esféricas (1,3 g / l).
-
Opción 2-900 μl de células EC (1 × 10 7 ) Se añadieron 100 μl de complejo híbrido (NP esféricas (1,3 g / l) + colesterol (0,55 g / l)).
El control fueron las células de la población total de CE, las cuales fueron incubadas en solución de glucosa al 5% sin tratamiento con nanocompuestos. El número de animales en cada grupo experimental no fue inferior a 20.
Después de la incubación, las células EC de todos los grupos probados se lavaron tres veces con solución salina (1:1) mediante centrifugación (10 min a 300 g).
La intensidad del desarrollo de CE después del pretratamiento con nanomateriales se evaluó mediante inyección intraperitoneal en una dosis de 3 × 10 6 células en 0,3 ml de solución salina. En 7 días después de la inoculación de células EC en todos los grupos estudiados, se determinó:
-
Un número total (TN) de células EC en la cavidad peritoneal.
-
Tasa de inhibición (Ri) del crecimiento de CE según la fórmula Ri =(TN (c) - TN (e)):TN (c) × 100%, donde TN (c) –– número total de células EAC en PC del grupo de control, TN (e):número total de células EAC en PC del grupo experimental.
-
La tasa de crecimiento (Rg) de EC se calculó usando la fórmula Rg (e) =Rg (c) - Ri, donde Rg (e) - tasa de crecimiento de tumores del grupo experimental de animales; Rg (c):tasa de crecimiento del tumor del grupo de control, Ri:tasa de inhibición del crecimiento de CE en un grupo experimental de animales; La tasa de inhibición del crecimiento de EC en el control se tomó como 100%, por lo que no hubo inhibición del crecimiento de EC.
-
CD44 alto / CD117 + proporción (proporción de CD44 alta porcentaje a CD117 + celdas).
-
La supervivencia de los animales se evaluó hasta el día 20 después de la inyección intraperitoneal de células EC no tratadas y tratadas con todo tipo de nanocompuestos.
El procesamiento estadístico se realizó utilizando Mann-Whitney U no paramétrico prueba en el software Statistica 6.0. Las diferencias se consideraron estadísticamente significativas en P <0.05.
Resultados
Los resultados obtenidos indican la presencia de una población heterogénea de células CE que portan en su superficie los marcadores CD44, CD24, Sca-1 y las que podrían atribuirse a elementos reguladores accesorios del microambiente (CD117). Las concentraciones de células con estas características en el grupo de CE total (grupo 1.1) se muestran en la Tabla 1 y son completamente consistentes con los hallazgos previos sobre la composición de la subpoblación de CE [28]. La identificación de la estructura Sca-1 virtualmente en todas las células EC permite considerarla como un marcador versátil de este tipo de tumor.
Lo más informativo en términos de identificación fenotípica de CSC es la expresión de la molécula CD44, que se usa por sí misma o en combinación con otros marcadores de superficie para aislar esta población celular de varios tumores, incluida la CE. Según las nociones clásicas, la diferenciación de las células tumorales durante el desarrollo del cáncer de mama se acompaña de la expresión reducida del receptor CD-44 con su desaparición gradual y aparición de las células que expresan el marcador CD24 [3].
Los candidatos para el papel de las CSC durante la EC podrían ser las células con CD44 alto fenotipo que forma parte de CD44 + CD24 - - población. Esta suposición sobre la dependencia de EC de la actividad funcional de una subpoblación de CD44 + Las células se probaron al evaluar la intensidad del crecimiento tumoral inducido por CD44 + y CD44 - facciones y población total de la CE. La Tabla 1 demuestra que la mayor actividad inductora de tumores era inherente a las células de CD44 + fracción. En realidad, después de administrar 3x10 6 CD44 + células (grupo 2.1), un número absoluto de células en PC fue 23 veces mayor que después de la población total de células EC (grupo 1.1), y 105 veces más que cuando CD44 - se administró fracción (Grupo 3.1).
Con esto, se encontraron cambios no solo de las composiciones cuantitativas sino también cualitativas de un tumor en desarrollo. La fracción de CD44 + formó la ascitis con un contenido predominante de CD44 + células, es decir, CD44 alto , CD44 + CD24 - y CD44 + CD24 + células. Además, la concentración de CD44 alta células fue 2 veces mayor en comparación con el grupo 1.1 y 16 veces mayor en el grupo 3.1. La fracción de CD44 - , por el contrario, formó un tumor que contenía más células maduras, es decir, aquellas con CD44 - CD24 + fenotipo. La misma redistribución de la composición de la subpoblación de células en el grupo 3.1 aparentemente determinó el contenido absoluto mínimo de células en la PC.
Es importante el hecho establecido de la presencia entre las células EC de una subpoblación con un CD117 + marcador. La molécula de CD117 es un receptor de tirosina quinasa transmembrana. En condiciones normales, es activado por el ligando correspondiente, es decir, el factor de crecimiento de células madre (SCGF) [29]. En patología oncológica se produce la activación dependiente de ligandona del receptor c-KIT, que en la mayoría de los casos (hasta un 92% de los casos) es consecuencia de la mutación del oncogén c-kit o está provocada por un mecanismo desordenado de regulación de la función de este receptor. [30].
Considerando CD117 + células como las células del microambiente tumoral el hecho establecido de la dependencia de la intensidad del crecimiento tumoral de la presencia o ausencia de CD117 + células y sus relaciones de concentración con CD44 alto células es lógica. Como muestra la Tabla 1, al iniciar la CE al introducir la población celular total (grupo 1.1.) En la PC, se formaron 34.80 ± 1.27 × 10 7 células en el CD44 alto / CD117 + ratio, que era igual a 0,02 unidades relativas.
Potencial tumorigénico de CD44 - fracción fue 4 veces menor (grupo 3.1) que se manifestó por un CD44 reducido alto / CD117 + proporción en la misma medida (4 veces) si se compara con el grupo 1.1. Este cambio en CD44 alto / CD117 + índice se debió principalmente a una disminución en CD44 alto concentración (en 8.5 veces) en el fondo del contenido reducido de CD117 + células (en 2 veces) también.
Al evaluar la intensidad del crecimiento de la ascitis, generada por CD44 + fracción, se observó un aumento significativo en el número total de células en PC (casi 24 veces si se compara con el grupo 1.1). Además, un factor importante es el exceso en dos veces de CD44 alto concentración y falta de CD117 + células. En material inicial de CD44 + fracción (antes del cultivo) según el análisis citométrico de flujo de los datos, el contenido de CD44 alto células fue 15 veces mayor que en la población total de células EC (datos no presentados).
Para resumir lo mencionado anteriormente, podemos argumentar que las CSC con una alta tasa de expresión del marcador CD44 (CD44 alto ) desempeñan un papel crucial en el inicio de la CE. ), mientras que una de las funciones más importantes de CD117 + La subpoblación es una regulación ("restricción") de la actividad tumorigénica de CD44 alta células. La ausencia de CD117 + células (grupo 2.1) parece multiplicar el potencial de diferenciación y proliferación de todo el grupo de CD44 + células, lo que provoca un aumento significativo en el número total de células en la PC.
El análisis del potencial proliferativo de un grupo total de células EC y CD44 + facción favorece esta interpretación. Se muestra que el factor de multiplicación (MF) en la población total cultivada en el CP en el grupo 2.1. durante 7 días aumentó casi en 24 veces si se compara con el grupo 1.1. Esto estuvo acompañado por una disminución en el tiempo celular que se duplicó de 24.47 ± 2.75 h en el grupo 1.1–14.70 ± 1.35 en el grupo 2.1 que puede caracterizar una población de células de ascitis cultivadas a partir de CD44 + fracción, como una de proliferación más activa (Tabla 1).
Para demostrar el papel especial de CD44 + células en el inicio y mantenimiento del tumor cuando se administra EC incluso en dosis mínimas, fue de interés evaluar comparativamente la capacidad tumorigénica de CD44 aislado + y CD44 - fracciones cuando se administra en diversas concentraciones. Se ha descubierto que después de la introducción de 3 × 10 6 células de la población de CE total, se observó crecimiento tumoral en el 100% de los animales (10/10) (Tabla 2). Reducing 10 times the dose of cells administered (3 × 10 5 ) resulted in a proportional decrease in absolute number of cells in the PC, tumor developed only in 50% of animals (Table 2). Reducing the administered dose of total EC population of cells down to 3 × 10 4 did not lead to tumor formation in the PC.
Initiations of EC by introducing of CD44 + cells at concentrations of 3 × 10 6 and 3 × 10 5 cells per animal resulted in almost 100% tumor development for both cases. Herewith, tumorigenic potential of CD44 + fraction exceeded that of total population of EC cells administered in the same doses (in 23 and 21 times, respectively). Moreover, introduction of 3 × 10 4 cells of CD44 + fraction caused a tumor formation in 33% of animals, while total population of EC cells used in the same dose, did not cause the formation of ascites. With the introduction of 3 × 10 3 cells of CD44 + fraction, no animals with the developed EC have been identified.
Fraction of CD44 – cells just in a dose of 3 × 10 6 was capable of forming tumors in 50% of animals, the number of cells in the PC in this case was 4.5 times less than when introducing the total population and in 105.9 times less than when inducing by CD44 + fraction. Thus, the results of this part of research suggest that CSCs are mainly present in the pool of cells with CD44 + phenotype. This emphasizes the importance of this subpopulation of cells in initiation and development of EC.
As noted above, identification and inactivation of CSCs is a major theoretical and practical issue of oncology. On this basis, the next task of our study was to investigate the impact of hybrid nanocomplexes designed at the Institute for Scintillation Materials of National Academy of Sciences of Ukraine on the tumorigenic activity of EC cells.
As Table 3 demonstrates an incubation of EC cells with only NPs as a component of hybrid nanocomplexes (option 1) decreased the concentration of CD44 high virtually twice if compared to the control and 5 times the content CD44 + CD24 – cells in ascites formed in vivo. The number in it of more differentiated CD44 + CD24 + , CD44 – CD24 + cells remained practically unchanged if compared to the control. In this group, there was established reduction of CD117 + cells (35%) at a slightly changed content of Sca-1 + subpopulation. Based on the data, the inhibition rate of EC growth (59.41 ± 3.45%) in variant 1 was accompanied by a twofold decrease in the concentrations of CD44 high cells in comparison with the control that was also reflected in the reduction of CD44 high /CD117 + ratio (Table. 3).
Pretreatment of EC cells with hybrid nanocomplexes (option 2) reduced almost 10 times the concentration of CD44 high and CD44 + CD24 – cells in the developed ascites if compared to the control (Table 3). It should be noted that the concentration of more differentiated CD44 + CD24 + and CD44 – CD24 + cells after this treatment increased slightly if compared to the control. The redistribution pattern of EC subpopulation composition in this option was accompanied with a pronounced enhancement of tumor growth inhibition compared to option 1 (74.70 ± 4.38 and 59.41 ± 3.45%, respectively, P < 0.05) that underlined the importance of cholesterol as a targeted compound of antitumor therapy. Pretreatment with hybrid nanocomplexes (option 2) led to maximal reduction there was found a maximum reduction of CD44 high /CD117 + ratio (10 times) as compared with option 1, that again confirmed a specific role of ratio of these cell subpopulations in the EC growth.
For all the types of EC pretreatment, the reduction of CD44 high /CD117 + ratio was accompanied by a decrease in tumor growth rate and increased survival of animals to day 20 of EC development (Fig. 3).

Tumor growth rate of EC, survival of animals and CD44 high /CD117 + ratio after incubation with nanocomplexes. Note:differences are statistically significant as compared with administration of the control (*), option 1 (**) (P < 0.05)
Discussion
One of the tasks of current oncology is elucidation of the mechanisms of initiation and development of malignant neoplasms. Mandatory participants in these events are the CSCs and so-called accessory-regulatory cells of tumor microenvironment. The variety of functional and structural characteristics of the CSCs in the development of different types of tumors determines the need for their further study. This is facilitated by the expansion of experimental model systems. One of them is the transplantable line of tumor cells of EC.
The elucidation of the peculiarities of this experimental model development, the subpopulation composition of tumor and tumorigenic potential of individual cell populations within the general pool of the EC cells will facilitate the development of new approaches to cancer therapy.
Using the method of phenotypic evaluation of progenitor cells of various levels of differentiation in the tumor focus makes it possible the identifying the stages, dynamics of development and invasiveness of the process. The established fact of heterogeneity of the EC subpopulation composition is important and there has been emphasized the value of CD44 + subpopulation in maintaining the growth of this type of tumor.
The most important role in implementing a tumorigenesis is played by an expression rate of the molecule. Indeed, in contrast to leukocytes for adhesion of those normally a low expression rate of CD44 receptor is required, triggering and self-maintenance in CSCs are implemented its much greater density on a cell surface [31].
It is known that CD44-glycoprotein is a hyaluronic acid (HA) receptor, a main component of extracellular matrix. The emerging set of HA-CD44 activates many receptor tyrosine kinases, resulting in activation of PI3K/Akt/ mTOR way [32, 33], which plays the role of a single universal signal transmission mechanism to the translation apparatus and is responsible for the integration of proliferative stimuli.
Among two known CD44-isoforms in normal hematopoietic cells its standard isoform (CD44s) is predominantly expressed [34]. In most malignant tissues there were detected both CD44s and variable isoforms of CD44- molecule (CD44v), resulting from alternative splicing of exons 6-15. Namely alternative splicing leads to a lengthening of CD44-extracellular domain, promoting its greater interaction with HA and tumor metastasis [35]. Due to that the role of CD44 high cells in triggering and maintaining the tumorogenesis is clear. It was previously found that a minor subpopulation of CD44 high cells had a high proliferative potential and played a critical role in EC developing [20].
In this paper, a special role of CD44 + -cells of the EC in initiation and maintenance of the tumor process in the EC under administration even in minimal doses has been shown. CD44 + cells were able to form a tumor even at a cell concentration of 100 times lower (10 4 cells/ mouse) if compared with the introduction of a total EC population (10 6 cells / mouse). The belonging of tumor cells to the CD44 + fraction was also confirmed by the fact that the EC initiation by the fraction of CD44-cells even at a dose of 10 6 cells / mouse caused the formation of a tumor only in 50% of cases, with an absolute number of cells in the PC 5 times less than in under introduction of a similar amount of the total population of EC and more than 100 times less than after the introduced CD44 + -fraction.
This is in accordance with the data of Shipitsin M et al. has shown that CD44 + and CD24 + cells in breast cancer development there are cell populations with different genetic profiles [36]. The research performed by Shipitsin M CD24 + cells have been noted to be more differentiated, while more progenitor-like functions are inherent to CD44 + células. The research performed by Shipitsin M CD24 + cells have been noted to be more differentiated, while more progenitor-like functions are inherent to CD44 + células. The authors suggest that CD24 + cells can be derived from CD44 + cells [36]. Fillmore C. and Kuperwasser C. supposed that CD24 + population was mainly characterized by less differentiated basal type of breast cancer, and CD44 + cells caused the development of luminal form of breast cancer, being more differentiated type of tumor [37].
Analyzing the patterns of tumor development, the classic hypothesis of «seed and soil» looks very actual [38], which postulated that an appropriate microenvironment (soil) is required for optimal growth of tumor cells (CSCs).
Most often the carcinoma-associated fibroblasts (CAFs) act as a tumor stroma in breast cancer and pancreatic cancer [39]. It has been shown that the CAFs, derived from invasive forms of human breast carcinomas, activated much stronger the growth of human breast cancer cell line MCF-7-Ras when administered to immunodeficient mice if compared with normal fibroblasts [9]. This function is implemented by the microenvironment cells due to the secretion by them of cytokines, chemokines and growth factors [10, 40].
Although so far the phenotypic identification of the microenvironment cells for various types of tumor has remained a subject of debate, most often used for this purpose the surface markers of primitive hematopoietic and endothelial cells, including c-kit (CD 117), CD133, VE-cadherin, VEGFR-2 and endoglin are used [41]. In this experimental model the most probable candidate to the role of tumor microenvironment cells is CD 117 + .
It is known that the c-KIT receptor (CD117 + ) is highly expressed in normal epithelium of the breast and progressively decreases with the development of breast carcinoma in situ and is almost completely lost in invasive breast cancer [42, 43]. Some authors proposed this kind of change in the expression rate of this marker as a possible test to assess the effectiveness of antitumor therapy [44].
Previously, after analysis of the significance of the content ratios for different subpopulations of EC cells when maintaining tumor growth, we proposed to use the CD44 high /CD117 + ratio as a prognostic criterion of tumor development [22].
Adequacy of using this index is confirmed in this study using the applied nanocomposites as therapeutic agents when treating the EC. The inhibition rate of EC growth (59.41 ± 3.45%) when treated with spherical NPs (option 1) was accompanied by a 2-fold decrease if compared to the control in the CD44 high -cell concentration, which was reflected in the reduced CD44 high / CD117 + index. The maximum decrease in the CD44 high /CD117 + index (10 times if compared to option 1) was established using the hybrid nanocomplexes for a pre-treatment of EC cells. Thus, many cells of a total pool of EC, but primarily those with the phenotype CD44 high and CD117 + , can be the target of the effect of the studied nanocomplexes (both direct and indirect). A significant decrease in their concentrations in the growing pool of EC after pretreatment with hybrid nanocomplexes clearly coincides with a reduced intensity of tumor growth.
Judging by the decrease in the amount of CD44 high as the most potent CSCs forming the entire subsequent series of advanced tumor cells, the main component in manifestation of antitumor effect of the synthesized hybrid nanocomplexes is spherical NPs. Introduction of cholesterol having affinity to tumor cell membranes into composition of hybrid nanocomplexes enhanced an inhibitory activity of NPs. Similar data were obtained by Betker J.L. et al. after analysis of the structure and functioning principles of the membranes of tumor cells. The authors concluded that the incorporation of cholesterol into membranes of tumor cells could be a prerequisite for a targeted delivery of liposomes with therapeutic agents directly into a cell.
Thus, the importance of cooperative interactions of cells with different phenotypic signs in maintaining the EC growth has been proven. The cells with the CD44 high phenotype being the part of the population of CD44 + CD24 – can be considered as CSCs in this model system. The use of new forms of nanocomposites that are capable to bind to CSCs and induce tumor destruction as the EC is a promising direction the treatment of oncopathology.
Conclusions
- 1.
On the base of the findings of phenotypic assessment and functional potential studies, the Ehrlich carcinoma is a heterogeneous population of tumor cells of varying differentiation extent referred to high and less potent tumor-inducing precursors, as well as the cells composing their microenvironment.
- 2.
A high (tenfold) tumorigenic activity of the EC CD44 + cells if compared to CD44 – cells was proven. In this pair of comparison, the CD44 + cells had a higher potential of generating in PC of CD44 high , CD44 + CD24 – , CD44 + CD24 + cell subpopulations, highlighting the presence of CSCs in a pool of CD44 + cells.
- 3.
There was found an ability of the synthesized nanocomplexes based on rare earth orthovanadates and cholesterol to inhibit the growth of CD44 + cell pool (CD44 high , CD44 + CD24 – , CD44 + CD24 + ) that was accompanied by a reduced intensity of EC growth (by 75%) and increased survival of the animal with tumors (in 3.5 times) in comparison with the control.
- 4.
It has been shown that the reduction in tumor growth rate after pretreatment with hybrid nanocomplexes was accompanied with a change in the composition of EC subpopulation that was reflected in a decrease in the CD44 high /CD117 + proporción. This ratio can be offered as one of diagnostic and prognostic tests of the severity and extent of oncology inactivation.
Abreviaturas
- BC:
-
Breast cancer
- CAFs:
-
Carcinoma-associated fibroblasts
- CSCs:
-
Cancer stem cells
- DiI:
-
1,1′-Dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine perchlorate
- EC:
-
Ehrlich carcinoma
- HA:
-
Hyaluronic acid
- MF:
-
Multiplicity factor
- NOD/SCID mice:
-
Nonobese diabetic-severe combined immunodeficiency mice
- NPs:
-
Nanopartículas
- PC:
-
Peritoneal cavity
- Rg:
-
Growth rate of EC
- Ri:
-
Inhibition rate of EC growth
- SCID mice:
-
Severe combined immunodeficiency mice
- TD:
-
Time doubling
Nanomateriales
- Baterías especiales
- Sangre artificial
- Célula solar
- Nanoárboles para células solares sensibilizadas con colorante
- Células solares de grafeno de alta eficiencia
- Nanoheterouniones para células solares
- Demostración de un biosensor flexible basado en grafeno para la detección rápida y sensible de células de cáncer de ovario
- Nanopartículas para la terapia del cáncer:avances y desafíos actuales
- Administración de medicamentos basados en células para aplicaciones de cáncer
- Un breve informe de progreso sobre las células solares de perovskita de alta eficiencia
- ¿Qué es un Stem Wall?