


El papel de la vía de la apoptosis en la citotoxicidad inducida por nanopartículas de óxido de zinc frescas y envejecidas
Resumen
Las nanopartículas de óxido de zinc (ZnO NP) se utilizan en una amplia gama de aplicaciones, incluida la industria, los productos comerciales y el campo de la medicina. Se realizaron numerosos estudios mecanicistas de la toxicidad de los NP de ZnO en NP prístinas (frescas). Sin embargo, la citotoxicidad inducida por las NP de ZnO transformadas (envejecidas) y los mecanismos subyacentes siguen sin estar claros. Aquí, observamos la transformación fisicoquímica de las NP de ZnO sufridas a lo largo del tiempo, seguida de la evaluación de la citotoxicidad de las NP frescas y envejecidas. Encontramos que las NP de ZnO frescas inducían un mayor nivel de apoptosis que sus contrapartes envejecidas. En consecuencia, los datos de secuenciación de ARN del híbrido humano-hámster tratado con ZnO NP envejecido ( A L ) mostraron que las vías de señalización p53, PI3k-Akt, FoXO, glutatión, ErbB, HIF-1, oxitocina y Jak-STAT estaban enriquecidas, pero ninguna vía de apoptosis. Los resultados de la PCR cuantitativa revelaron un nivel de ARNm significativamente más alto de IL1B y CD69 en grupos tratados con NP fresco en comparación con los grupos tratados con NP de ZnO y cloruro de zinc envejecidos. Los resultados anteriores indicaron que la menor citotoxicidad de las NP de ZnO envejecidas se atribuye parcialmente a su potencia reducida para inducir la apoptosis. La regulación transcripcional de múltiples vías de señal activadas por NP envejecidas puede ayudar a construir la homeostasis celular. Tomados en conjunto, nuestros hallazgos destacan la influencia del proceso de envejecimiento (transformación ambiental) de los NP de ZnO en sus toxicidades y consecuencias biológicas.
Introducción
Con el rápido desarrollo de la nanotecnología en las últimas décadas, las nanopartículas (NP) se han aplicado en varios campos, incluida la industria, la vida diaria humana y la nanomedicina [1, 2]. El Inventario de productos de consumo de nanotecnología (CPI) muestra un aumento de 30 veces entre 2005 y 2015 en el número de nanoproductos, incluidos 762 productos de salud (fitness), 72 alimentos (bebidas) y 23 productos para bebés [2]. La creciente aplicación de NP en productos de consumo y en varios campos aumentó la posibilidad de que las NP ingresen al medio ambiente, lo que genera preocupaciones de seguridad con respecto a sus posibles impactos adversos. Los NP de óxido de zinc (ZnO) se encuentran entre los NP más utilizados y su producción anual mundial ha alcanzado casi las 3400 toneladas [3, 4]. Algunas sustancias que antes se consideraban biológicamente inertes podrían volverse tóxicas en su estado de nanopartículas. Un número cada vez mayor de estudios ha aclarado que las NP de ZnO pueden plantear riesgos importantes para las células de mamíferos y los animales al inducir una toxicidad significativa [5, 6, 7].
Se han utilizado diversas estrategias que incluyen el recubrimiento, la funcionalización de la superficie y la modificación del estado de oxidación para atenuar la toxicidad potencial de las NP modificando sus propiedades físicas y químicas (como la disolución, aglomeración y perturbación de las membranas celulares) [8,9,10, 11]. Aunque estas modificaciones de los NP debilitan sus efectos tóxicos en cierto grado, los usos de los NP no siempre son seguros, especialmente en determinadas condiciones y entornos de exposición [12,13,14]. En realidad, muchos tipos de NP no son estables y tienden a sufrir un "envejecimiento" o una "transformación ambiental" después de haber sido liberados intencionalmente o no en el ambiente natural [14, 15, 16, 17]. En los últimos años se ha trabajado mucho para explorar el proceso de transformación ambiental de las PN; sin embargo, la investigación sobre los efectos tóxicos de los NP "transformados (envejecidos)" es todavía muy limitada, por no hablar de sus mecanismos tóxicos.
Como representante típico de las NP no persistentes, las NP de ZnO tienen una reactividad muy alta y son propensas a transformarse en propiedades físicas y químicas y en un estado de aparición después de ser liberadas al medio ambiente o ingeridas por animales, lo que podría afectar significativamente sus efectos toxicológicos [17 , 18]. Por ejemplo, los estudios han demostrado que el proceso de sulfuración de las NP de ZnO cambió la carga, la hidrofobicidad y el estado de agregación, lo que resultó en la adsorción de las NP en estado sulfuro en la saliva humana, el sudor y el líquido de lavado broncoalveolar. Además, la proteína adsorbida por ZnO NPs forma una corona proteica especial que generalmente afecta su efecto biológico [19]. Los fosfatos en soluciones fisiológicas podrían convertir las NP de ZnO en ZnHPO 4 metaestable y Zn 3 (PO 4 ) 2 en aproximadamente 5 a 10 h [20]. El proceso de transformación completa de NP de ZnO (≤ 3 μg / mL) en el sistema de exposición in vitro a linfocitos T humanos (37 ° C, medio de cultivo celular RPMI1640 que contiene 10% de FBS durante 24 h) se investigó utilizando radiación de sincrotrón X- espectroscopia de estructura cercana al borde de absorción de rayos (XANES) [21]. Los estudios anteriores sugieren la subestimación de los riesgos ambientales y para la salud de las NP de ZnO al evaluar únicamente los efectos biológicos de las NP de ZnO prístinas (frescas). A la luz de este problema, existe una necesidad urgente de comprender de manera integral los procesos de envejecimiento y transformación ambiental de las NP [22].
Nuestro estudio anterior reveló que las NP de ZnO envejecidas durante 40 a 120 días en agua ultrapura se someten a una transformación fisicoquímica y se convierten en Zn 5 (CO 3 ) 2 (OH) 6 , Zn (OH) 2 y Zn 2+ [23]. Curiosamente, las NP de ZnO envejecidas exhibieron una citotoxicidad más baja que las contrapartes frescas [23], sin embargo, los mecanismos de toxicidad de este tipo de variación no están claros. En el presente estudio, nos propusimos explorar las razones subyacentes de las diferentes citotoxicidades entre NP de ZnO frescas y envejecidas. Se aplicaron sistemáticamente NP de ZnO con dos tamaños de partículas diferentes (20 nm y 90-200 nm). Los ensayos de citotoxicidad demostraron que las NP de ZnO envejecidas inducían anomalías morfológicas menos pronunciadas y viabilidades celulares relativamente más altas que sus contrapartes frescas. Los datos de secuenciación de ARN revelaron que los genes apoptóticos se enriquecieron en células frescas tratadas con ZnO NP, mientras que estos genes se vieron mucho menos afectados por los tratamientos envejecidos con ZnO NP. Además, las células expuestas a NP de ZnO envejecidas mostraron un nivel reducido de proteína Caspasa-3 escindida, lo que indica además la mayor potencia de las NP de ZnO frescas para provocar la apoptosis en células cultivadas. En combinación con nuestros hallazgos anteriores, este estudio sugirió que la citotoxicidad disminuida de las NP de ZnO envejecidas se atribuye a su capacidad atenuada para desencadenar la apoptosis celular.
Materiales y métodos
Nanopartículas y reactivos
Los nanopolvos de ZnO disponibles comercialmente (ZnO NP), con un tamaño promedio informado por el fabricante de 20 nm (99,5% de pureza, casi esférico) y 90-200 nm (99,9% de pureza, morfología irregular), se compraron en Nanotructured &Amorphous Materials (Houston, TX ). Salvo que se indique lo contrario, todos los reactivos y productos químicos utilizados en este estudio se compraron a Sigma-Aldrich (Shanghai, China).
Dispersión, envejecimiento y caracterización de nanopartículas
Se prepararon suspensiones madre de ZnO NP (1 mg / ml) suspendiendo nanopolvos secos en Milli- Q H 2 O (Millipore, 18 MΩ cm) y esterilizado en autoclave (120 ° C, 30 min) y luego almacenado a 25 ° C durante un período de envejecimiento natural que varía de 0 a 60 días. Las NP de ZnO transformadas de forma natural de 0 y 60 días se designaron NP frescas y envejecidas, respectivamente. Para asegurar una dispersión adecuada, las suspensiones frescas y envejecidas se sonicaron (100 W) durante 30 min en un baño ultrasónico antes de la caracterización o incubación con células. La morfología, el tamaño de partícula y la agregación de NP de ZnO frescas y envejecidas se caracterizaron mediante microscopía electrónica de transmisión (TEM, JEOL JEM-2010, Tokio, Japón). La estructura cristalina de NP de ZnO frescas y envejecidas se determinó usando difracción de rayos X de potencia (XRD, PANalytical B. V., Shanghai, China) comparando con estándares auténticos. Los detalles del proceso de envejecimiento natural y la caracterización de las NP de ZnO se han descrito anteriormente [23].
Cultivo celular y tratamiento con NP de ZnO
A L línea celular, un tipo de células híbridas humano-hámster formadas por la fusión de gly2A En este estudio se utilizó mutante de ovario de hámster chino (CHO) y fibroblastos humanos. Estas células híbridas contenían un conjunto estándar de cromosomas CHO-K1 y una única copia del cromosoma humano 11 y se cultivaron en medio Ham's F12 (Hyclone, Grand Island, NY) suplementado con suero fetal bovino (8%, Hyclone, Grand Island, NY) ), gentamicina (25 g / mL) y glicina (2 × 10 –4 M) a 37 ° C en un 5% CO 2 humidificado incubadora [24]. Las suspensiones madre de NP de ZnO frescas y envejecidas se dispersaron mediante 30 min de ultrasonidos (100 W) para evitar la aglomeración, posteriormente se diluyeron a concentraciones apropiadas con medio de cultivo celular para la exposición de las células. Las células mantenidas en medios de cultivo celular sin NP sirvieron como control en cada experimento.
Ensayo para detectar la citotoxicidad
A L Las células en una fase logarítmica de crecimiento se cultivaron en portaobjetos de vidrio en placas de Petri de 35 mm (6 × 10 4 células / placa) durante 24 h antes de la estimulación, seguido de tratamiento con 2 mL de medio de cultivo que contiene 1, 5, 10, 12, 15 y 20 µg / mL NP de ZnO fresco o envejecido 72 h. Una vez finalizado el tiempo de tratamiento, las imágenes de la morfología celular se obtuvieron utilizando un microscopio Leica DM4B (Leica, Alemania). ZnCl 2 se incluyó como referencia de iones de zinc para comparar la citotoxicidad con NP de ZnO.
Se utilizó el kit de recuento celular (CCK-8) (APExBIO, Shanghai, China) para detectar la viabilidad celular. En detalles, A L las células se sembraron en placas de 96 pocillos (4 × 10 3 células / pocillo) con medio de cultivo celular durante 24 hy tratado con medio que contiene varias concentraciones de ZnCl 2 , NP de ZnO fresco y envejecido durante 24, 48 y 72 h, respectivamente. Para la solución de trabajo, el volumen de NP añadidas de la suspensión madre fue menos del 5% del volumen total del medio de cultivo en cada pocillo. Una vez finalizado el tiempo de tratamiento, se aspiró el medio de cultivo y las células se incubaron con 100 µL de solución de trabajo CCK-8 durante 2 ha 37 ° C siguiendo las instrucciones del fabricante. Luego, se registró la absorbancia a 450 nm usando un lector de fluorescencia Spectra Max M2 (Molecular Devices, Wokingham, Berks, Reino Unido). La viabilidad celular se calculó como un porcentaje de absorbancia en los pocillos, con cada concentración de NP normalizada a la absorbancia de las células de control (100%).
Extracción de ARN, transcripción inversa y PCR cuantitativa
A L Las células en una fase logarítmica de crecimiento se sembraron en una placa de Petri de 35 mm de diámetro (6 × 10 4 células / placa) con medio de cultivo celular durante 24 h. Luego, el medio se reemplazó con 2 mL de medio de cultivo que contenía 12 µg / mL de ZnCl 2 , NP de ZnO fresco y añejado durante 72 h. Una vez finalizado el tiempo de tratamiento, se aspiró el medio de cultivo y las células se lavaron 3 veces con PBS. Posteriormente, se añadió 1 ml de reactivo Trizol (Invitrogen, Carlsbad, CA, EE. UU.) A cada placa para extraer el ARN total de acuerdo con las instrucciones del fabricante. La concentración y pureza del ARN total obtenido después de la extracción se cuantificó utilizando un espectrofotómetro Q5000UV-Vis (Quawell, EE. UU.). Después de la cuantificación, se realizó la transcripción inversa utilizando el kit TransGene RT-PCR (TransGene Biotech, Beijing, China) para obtener el ADNc de la plantilla de ARN de acuerdo con los protocolos del fabricante. Las muestras de ADNc resultantes se cuantificaron utilizando el espectrofotómetro UV-Vis Q5000 y luego se analizaron utilizando SYBR-Green como tinte de fluorescencia (TransGene Biotech, Beijing, China) en el sistema Roche RT-PCR (Applied Biosystems) [25].
El gen de mantenimiento que codifica la gliceraldehído-3-fosfato deshidrogenasa ( Gapdh ) se utilizó como control interno para evaluar Il-1α , Il-1β , Caspasa 3 , CD69 , junio y MT1 Expresión de ARNm. Los resultados se expresaron como la relación de expresión relativa entre el gen objetivo y Gapdh . Las secuencias de cebadores utilizadas en este estudio se proporcionan en la Tabla 1.
Análisis de datos de secuenciación de ARN
Las muestras de ARN total de A L células del grupo de control, grupo tratado con NP de ZnO envejecido y ZnCl 2 grupo tratado fueron secuenciados por BangFei Bioscience (Beijing, China). Brevemente, el ARN total de A L Las células se extrajeron siguiendo los protocolos de TRIZOL, hasta la precipitación isoproponal. Luego, las muestras de ARN se resuspendieron en el tampón de extracción antes de la secuenciación. Los datos de secuenciación de ARN de recuento sin procesar se analizaron utilizando el paquete R Deseq2 [Eric1]. El diagrama de Venn fue generado por el paquete R VennDiagram [Eric1.2]. Los genes significativamente modificados se utilizaron para un análisis adicional de enriquecimiento de la vía. Los experimentos se realizaron en tres repeticiones independientes. Se excluyeron del análisis los genes de ARNr, los genes mitocondriales y los genes detectados con menos de 40 pb.
Los datos de secuenciación de ARN, series de referencia GSE97852, GSE60159 y GSE39444, se obtuvieron de Gene Expression Omnibus [Eric 2, 3, 4]. R (versión 3.6.2) generó el gráfico de análisis de enriquecimiento del conjunto de genes utilizando el paquete fgsea [Eric 5]. Los genes de la apoptosis con un cambio significativo de 1,5 veces & p Se utilizaron valores <0,05 para análisis adicionales. El mapa de calor con árbol de genes fue generado por el paquete R "ComplexHeatmap" [Eric 6]. La vinculación promedio se usó como método de agrupamiento, y Euclidean se usó como método de medición de distancia. El análisis de enriquecimiento de la vía fue precedido por STRING2.0 [Eric 7].
Western Blot
A L Las células en una fase logarítmica de crecimiento se sembraron en una placa de Petri de 60 mm de diámetro (1,5 × 10 5 células / placa) con medio de cultivo celular durante 24 h. Luego, el medio se reemplazó con 4 mL de medio de cultivo que contenía 12 µg / mL de NP de ZnO frescas o envejecidas durante 24 h. Al final del período de exposición, se aspiró el medio de cultivo y luego, las células se lavaron 3 veces con PBS y se lisaron en hielo con tampón de lisis RIPA (Beyotime, China) para recolectar proteínas celulares. Se separaron cantidades iguales de proteínas celulares en geles SDS-PAGE al 12% y luego se transfirieron a una membrana de fluoruro de polivinilideno (PVDF) (Roche, Suiza). Brevemente, después de 2 h de bloqueo con leche desnatada al 5% en TBST a 25 ° C, las membranas se incubaron posteriormente con anticuerpo primario en diluciones apropiadas (de acuerdo con los protocolos del fabricante) a 4 ° C durante la noche, seguido de incubación con secundario conjugado con HRP. anticuerpos (1:5000, Promega, Madison, EE. UU.) durante 2 ha 25ºC. Finalmente, el inmunomarcado se detectó usando una solución de quimioluminiscencia mejorada (ECL) (BOSTER, China). Los anticuerpos primarios de anti-pro / caspasa-3 escindida y anti-actina se adquirieron de Cell Signaling Technology e ImmunoWay, respectivamente.
Estadísticas
El análisis estadístico se compiló sobre la media de los resultados obtenidos de al menos tres experimentos independientes. Todos los datos se presentaron como medias ± desviación estándar (DE) y se compararon estadísticamente mediante el análisis de varianza unidireccional (ANOVA). En todas las parcelas p los valores <0.05 se mostraron como * y se consideraron estadísticamente significativos.
Resultados
Caracterización de NP de ZnO
Para determinar las diferencias en las características fisicoquímicas detalladas entre las NP de ZnO frescas y envejecidas, primero observamos la morfología de las NP utilizando TEM (Fig. 1A). Nuestros resultados indicaron que las NP de ZnO frescas de 20 nm eran cristales casi esféricos y las NP de ZnO frescas de 90-200 nm eran cristales cúbicos / en forma de varilla irregularmente. El tamaño de partícula individual fue consistente con el tamaño proporcionado por el fabricante. Obviamente, tanto las NP de ZnO de 20 nm como las de 90–200 nm estaban inclinadas a agregarse en agua ultrapura. Además, independientemente de la forma y el tamaño de las NP originales, la microestructura de las NP de ZnO de 20 nm y 90-200 nm cambió drásticamente de una estructura cristalina transparente a un estado amorfo o en forma de hoja / aguja después de envejecer durante 60 días. Además, la naturaleza cristalina y la pureza de fase de las NP frescas y envejecidas se determinaron mediante difracción de rayos X (XRD) con radiación Cu Kα ( λ =0,15418 nm) se acercan a 25 ° C, como se muestra en la Fig. 1B. El patrón XRD de NP de ZnO frescas indicó que las muestras estaban compuestas por una estructura de wurtzita cristalina y no se identificaron picos de impureza característicos, lo que sugiere una alta calidad de NP frescas. Para los NP envejecidos, el patrón XRD exhibió la neoformación de Zn 5 (CO 3 ) 2 (OH) 6 (número de tarjeta 00-011-0287) y Zn (OH) 2 (número de tarjeta 00-003-0888) fases sólidas, que indican la transformación química de las NP de ZnO (20 y 90-200 nm) durante el proceso de envejecimiento.
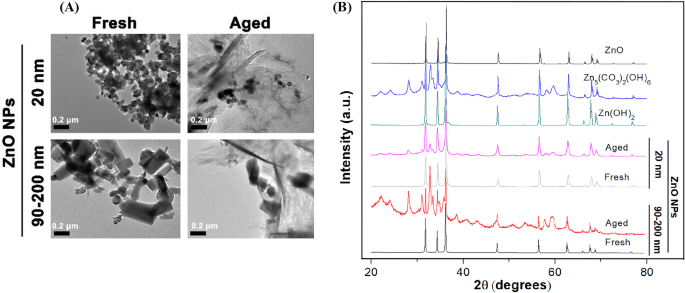
Características fisicoquímicas de NP de ZnO frescas y envejecidas. A Micrografías representativas de NP frescas y envejecidas (100 μg / mL, 20 y 90-200 nm) en Milli- Q agua usando TEM de baja resolución, B Patrones XRD de NP frescos, NP envejecidos, ZnO, Zn (OH) 2 y Zn 5 (CO 3 ) 2 (OH) 6 referencias en forma seca
Observación morfológica de A L Células expuestas a NP de ZnO frescas y envejecidas
El tratamiento de los NP da como resultado un cambio notable en la forma o morfología celular in vitro [26]. Por lo tanto, A L las células expuestas a NP de ZnO frescas o envejecidas a 10 y 15 µg / ml durante 72 h se examinaron bajo un microscopio estereoscópico. Como se muestra en la Fig. 2, la morfología celular en el grupo de control permaneció normal. Las células se adhirieron bien y la mayoría se adhirió en 2 h. La mayoría de las células tenían forma de huso o poligonales, y algunas células recién divididas mostraban un citoplasma más transparente y una mejor dispersión durante el proceso de adhesión. El tratamiento con NP de ZnO fresco de 12 μg / ml (20 nm y 90–200 nm) durante 72 h cambió significativamente la morfología celular. Aunque la mayoría de las células se adhirieron en 3-5 h, no pudieron diseminarse bien y algunas células se redondearon y perdieron la forma poligonal. Cuando la concentración de ZnO NP se incrementó a 15 μg / mL, las células tratadas se atrofiaron y no pudieron adherirse, lo que sugiere su viabilidad celular significativamente menor que la de las células tratadas con 10 μg / mL. Estos resultados indicaron que la LC100 para NP de ZnO frescas es probablemente menor de 15 μg / mL por un tratamiento de 72 h. Por el contrario, la morfología celular en los grupos tratados con NP de 20 nm y 90-200 nm de edad (15 μg / ml) no se vio significativamente afectada, y la mayoría de las células supervivientes pudieron adherirse y diseminarse, observándose menos de la mitad de las células muertas, lo que revela que Los NP de ZnO envejecidos son mucho menos citotóxicos que los NP de ZnO frescos.
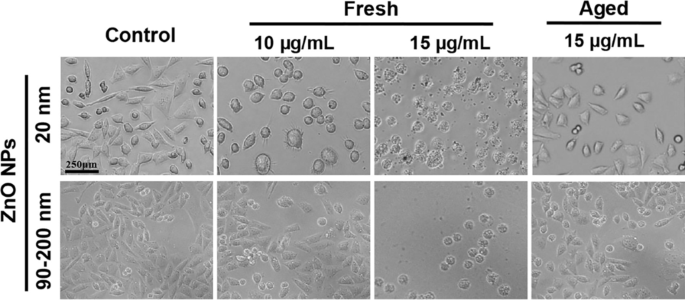
Cambios morfológicos en A L células después de la exposición a NP de ZnO frescas o envejecidas durante 72 h en medio F12 de Ham suplementario, y las células no expuestas se utilizaron como grupos de control. A L La morfología celular se observó con un microscopio óptico con un aumento de 10 ×
Los NP de ZnO envejecidos indujeron una menor citotoxicidad que los NP frescos
Para investigar más a fondo la diferencia en la citotoxicidad entre NP de ZnO frescas y envejecidas, examinamos la viabilidad celular mediante el uso de kits CCK-8. Como se muestra en la Fig.3, incubación A L las células con dosis en gradiente de NP de ZnO frescas y envejecidas (de 0 a 20 μg / ml, 20 nm y 90-200 nm) durante 24 h, 48 ho 72 h mostraron una disminución de la viabilidad celular dependiente de la dosis. No se observó citotoxicidad obvia al tratar las células con NP de ZnO ≤ 10 μg / ml. Cuando la dosis de NP de ZnO frescas y envejecidas se elevó a 12 y 15 μg / mL, la viabilidad celular mostró una tendencia a la disminución dependiente del tiempo. Obviamente, la viabilidad celular en los grupos tratados con NP de edad avanzada fue significativamente mayor que en los grupos tratados con NP reciente. Además, ZnCl 2 -tratamiento también comprometió la viabilidad celular de una manera dependiente de la dosis y el tiempo, mientras que la citotoxicidad de ZnCl 2 fue mucho menor que la de los NP de ZnO frescos y envejecidos.
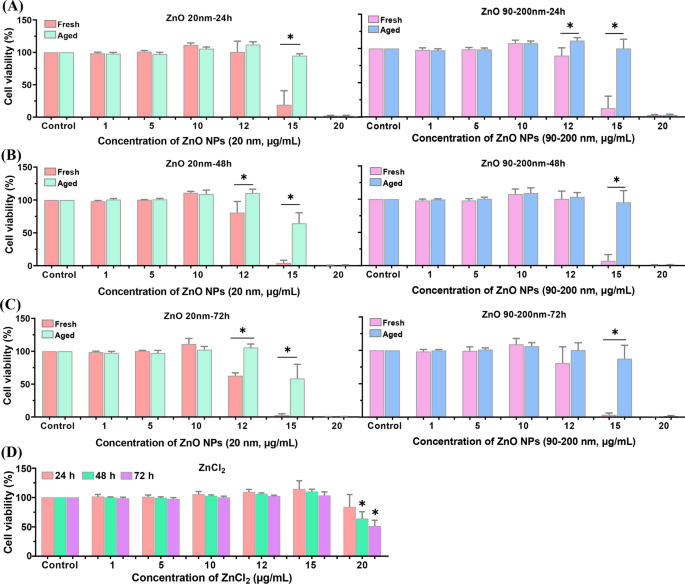
Viabilidad celular inducida por NP de ZnO frescas y envejecidas en A L células. A L Las células se incubaron con diversas concentraciones de NP de ZnO frescas y envejecidas (20 y 90-200 nm) durante 24 h ( A ), 48 h ( B ) y 72 h ( C ). D A L las células fueron expuestas a varias concentraciones de ZnCl 2 para diferentes momentos. Los datos se basaron en ≥ 3 experimentos independientes y se expresaron como media ± DE, * p <0.05
El tratamiento de NP de ZnO fresco activó las vías de apoptosis y aumentó la expresión de genes apoptóticos
Para develar los mecanismos subyacentes que conducen a la menor citotoxicidad de las NP envejecidas, analizamos los datos de secuenciación de ARN de las NP de ZnO frescas y envejecidas. Como se muestra en la Fig. 4A, B, después del tratamiento con NP de ZnO frescas, se activó la vía de apoptosis en las células Jurkat ( p =0.017) y celdas HMDM ( p =0,041). Los genes de la apoptosis: ANXA1 , CYLD , TNFSF10 , IER3 , CDKN1A , JUN , SAT1 , PMAIP1 , CD38 y ISG20 se enriquecieron significativamente en células Jurkat frescas tratadas con ZnO NP. Los genes de la apoptosis: CD38 , TNFRSF12A , CCNA1 , BMP2 , PPP2R5B , EREG , IFNGR1 , CD44 , CD14 , GNA15 , GCH1 , TIMP1 , BTG2 , IL1B , IL1A , BTG3 , BCL2L11 , SC5D y SPTAN1 se enriquecieron significativamente en células HMDM frescas tratadas con ZnO NP (Fig. 4C, D). Dado que las células de Jurket (células de linfocitos T de sangre periférica) y las células HMDM (macrófagos derivados de monocitos humanos) son diferentes tipos de células, la forma en que desencadenan la apoptosis puede ser diferente. En resumen, estos resultados mostraron que la exposición fresca de NP de ZnO podría activar diferentes vías de apoptosis en varios tipos de células.
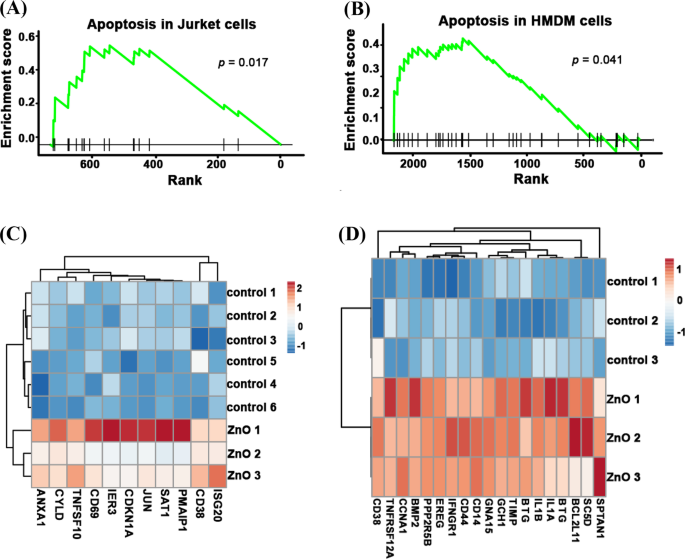
La ruta de la apoptosis se enriqueció en datos de secuencia de ARN de células Jurket y HMDM nuevas tratadas con ZnO NP. La puntuación de enriquecimiento de genes expresados significativamente de la vía de apoptosis de células Jurket frescas tratadas con ZnO NP ( A ) y celdas HMDM ( B ). El mapa de calor de la expresión de genes apoptóticos de células Jurket frescas tratadas con ZnO NP ( C ) y celdas HMDM ( D ) y sus grupos de control
Los NP de ZnO envejecidos no regularon positivamente la expresión de genes apoptóticos como NP de ZnO frescos
Nuestros datos de secuenciación de ARN de A tratados con NP de ZnO envejecido L las células mostraron que las vías de señalización p53, PI3k-Akt, FoXO, glutatión, ErbB, HIF-1, oxitocina y Jak-STAT estaban enriquecidas (Fig. 5A). Los genes de apoptosis enriquecidos en células Jurket y HMDM no se vieron afectados significativamente en las células envejecidas tratadas con ZnO NP (Fig. 5B). Para confirmar aún más los hallazgos, probamos la expresión de genes relacionados mediante PCR en tiempo real. Encontramos que algunos de los genes de apoptosis: BMP2 , PMAIP1 , IL1α , CD69 , CCNA1 , CD38 y IL1β eran indetectables en A tratadas con NP de ZnO envejecidas L células (datos no mostrados), probablemente porque la mayoría de estos genes se expresan en células del sistema inmunológico. Los otros genes de apoptosis regulados positivamente ( IL1α , IL1β y CD59 ) observados en grupos tratados con NP de ZnO frescos no cambiaron significativamente en los niveles de expresión por el tratamiento de NP de ZnO envejecidas. Mientras que el MT1 que sirven como control positivo se incrementó significativamente en un nivel de expresión, la expresión de Caspase 3 no se modificó significativamente (Fig. 5C). Estos datos sugirieron que las NP de ZnO envejecidas, a diferencia de sus contrapartes frescas, son menos potentes para activar los genes de la vía de la apoptosis en A L células.
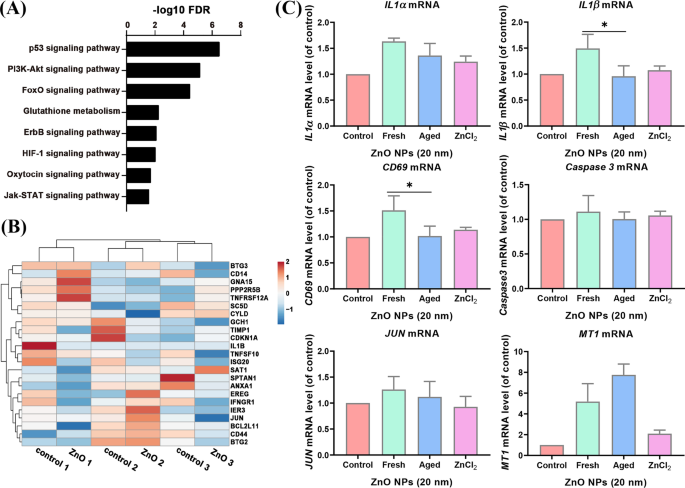
La vía de la apoptosis no se enriqueció en los datos de la secuencia de ARN de A tratado con NP de ZnO envejecido L células. ( A ) El análisis de ontología genética de vías enriquecidas de A tratadas con NP de ZnO envejecido L células. ( B ) El mapa de calor de la expresión génica apoptótica de A tratada con NP de ZnO envejecida L células y grupo de control. ( C ) La expresión de genes apoptóticos seleccionados y genes de control ( MT1 ) en A tratado con NP de ZnO fresco y envejecido L celdas
Los NP de ZnO frescos pero no envejecidos aumentaron el nivel de expresión de la proteína caspasa 3 activada
Detección de la expresión génica de caspasa 3 por sí solo no puede indicar directamente la activación de la vía de la apoptosis. Para analizar más a fondo si el tratamiento con ZnO NP podría cambiar el nivel de proteínas apoptóticas, se examinó la expresión de la proteína caspasa 3 escindida, un biomarcador de uso común para indicar la activación de la apoptosis celular [27], mediante análisis de transferencia Western. Como se muestra en la Fig.6, en comparación con el grupo de control, el tratamiento con NP de ZnO fresco (20 nm) aumentó el nivel celular de la proteína Caspasa 3 escindida en 1,31 ± 0,023 veces, que fue significativamente más alto que el de NP de ZnO de 20 nm de edad. grupo tratado (1,12 ± 0,039 veces). Cuando el tamaño de partícula de las NP frescas de ZnO se incrementó a 90-200 nm, la expresión de la proteína caspasa 3 escindida inducida por NP frescas aumentó 1,46 ± 0,078 veces, significativamente mayor que la de las NP envejecidas (1,07 ± 0,075 veces) . Estos datos ilustraron aún más la mayor potencia de las NP de ZnO frescas para inducir la apoptosis celular, en comparación con sus contrapartes envejecidas.
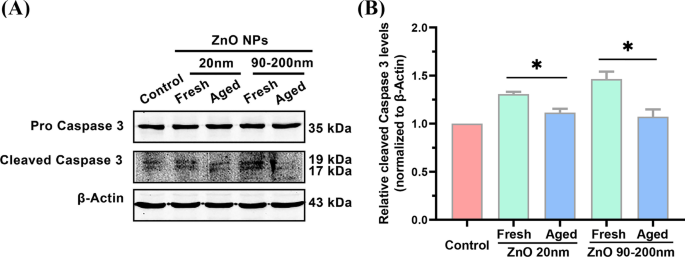
Niveles apoptóticos en A L células incubadas con NP de ZnO frescas y envejecidas (20 y 90–200 nm). Análisis de Western Blot ( A ) y cuantificación ( B ) de los niveles de proteína caspasa 3 escindida cuando las células se incubaron con NP de ZnO frescas y envejecidas de 12 μg / ml (20 y 90-200 nm) durante 72 h. Los datos se basaron en ≥ 3 experimentos independientes y se expresaron como media ± DE, * p <0.05
Discusión
Se informó que las NP de ZnO experimentan una transformación fisicoquímica en Zn 5 (CO 3 ) 2 (OH) 6 con el lanzamiento de Zn 2+ durante el proceso de envejecimiento natural [23, 28]. Sin embargo, la citotoxicidad inducida por las NP de ZnO transformadas (envejecidas) y los mecanismos subyacentes siguen sin estar claros. Aquí, para develar el mecanismo de citotoxicidad diversa entre NP de ZnO frescas y envejecidas, se llevaron a cabo análisis de secuenciación de ARN y prueba de RT-PCR. Además, se aplicó transferencia de Western para examinar el nivel de proteína de Caspasa 3, el ejecutor clave en la apoptosis celular.
Nuestros datos mostraron que las NP de ZnO envejecidas inducían mucha menos citotoxicidad que las NP de ZnO frescas en A L células. El LC 100 de ambas NP de ZnO frescas (90-200 nm y 20 nm) en nuestro estudio actual fue inferior a 15 μg / ml (Fig. 3), lo cual fue consistente con hallazgos previos de que la LC 100 de NP de ZnO con 19-36 nm para células NIH-3T3 o MSTO es de aproximadamente 15 μg / ml [29]. Confirmamos que las transformaciones ambientales de las propiedades fisicoquímicas de las NP pueden alterar drásticamente su toxicidad. Se ha informado que el proceso de sulfuración de las NP de ZnO cambia su carga, hidrofobicidad y estado de agregación, dando como resultado la adsorción de NP en estado sulfuro en la saliva humana, el sudor y el líquido de lavado broncoalveolar. Y la proteína adsorbida por ZnO NPs formó una corona de proteína especial, que afectó su efecto biológico [19]. Los fosfatos ampliamente presentes en soluciones fisiológicas (como la saliva) podrían convertir las NP de ZnO en ZnHPO 4 metaestable y Zn 3 (PO 4 ) 2 dentro de aproximadamente 5 a 10 hy mostró citotoxicidad para las células epiteliales del tracto digestivo [20]. Ivask y col. demostró la ocurrencia de la transformación completa de NP de ZnO (≤ 3 μg / mL) en el sistema de exposición in vitro a linfocitos T humanos (37 ° C, medio de cultivo celular RPMI1640 que contiene 10% FBS durante 24 h) usando radiación de sincrotrón absorción de rayos X espectroscopía de estructuras cercanas al borde (XANES). El espectro y la citotoxicidad de los productos de transformación fueron consistentes con los de ZnSO 4 [21]. Nuestros resultados revelaron la toxicidad dependiente de la dosis y el tiempo de ZnCl 2 a A L células, donde su citotoxicidad es mucho menor que las NP de ZnO frescas y envejecidas (Fig. 3). La observación explica además el hallazgo de que la citotoxicidad de ZnO NP fresco no se atribuye completamente a su Zn 2+ liberado [30].
Nuestro estudio anterior también mostró que las NP de ZnO envejecidas exhiben una mayor potencia para provocar ROS (especies reactivas de oxígeno), así como una capacidad atenuada para matar células en comparación con las NP de ZnO frescas [23]. Razonamos que la menor citotoxicidad inducida por NP de ZnO envejecidas podría ser más tolerable en células de mamíferos. El presente estudio de los datos de secuenciación de ARN ilustró que los genes apoptóticos se han regulado positivamente en células frescas tratadas con ZnO NP, donde se vieron mucho menos afectados en los grupos tratados con NP de edad avanzada. IL1α e IL1β son miembros de la familia de citocinas de la interleucina 1. La liberación de IL1α e IL1β activa la apoptosis parcialmente dependiente de Caspasa 8 [31]. CD69 codifica un miembro de la superfamilia de receptores transmembrana de tipo II de lectina dependiente de calcio. El aumento de la expresión de CD69 se asoció con un aumento de la expresión de la apoptosis anexina V y el marcador CD95 (Fas) [32]. JUN es una subunidad del factor de transcripción AP-1. El aumento de la actividad de JUN escinde proteolíticamente la alfa-fodrina, un sustrato de la enzima convertidora de interleucina 1beta (ICE), y la familia CED-3 de cisteína proteasas, que además provoca la muerte celular programada [33]. El aumento de la expresión de estos genes apoptóticos reveló que las NP recientes desencadenan la apoptosis de varias formas diferentes. After the elevation of these apoptotic gene expressions, apoptosis processes are eventually executed by apoptotic proteins (Fig. 7). Caspase 3 is the core protease for various apoptotic scenarios; cleavage of this protein is necessary to activate both extrinsic and intrinsic apoptotic pathways [34, 35]. Therefore, detection of cleaved caspase 3 is a common method for identifying apoptosis induced by a wide variety of apoptotic signals [36]. Our Western blotting data revealed that, for both 20 nm and 90–200 nm ZnO NPs, sublethal exposure did not alter the level of Pro caspase 3 in all treatment groups. In contrast, cleaved Caspase 3 was significantly elevated by fresh NPs treatment, where aged NPs showed few (if any) effects on the level of cleaved caspase 3 (Fig. 6). Combined with RNA expression analysis, our results clearly elucidated the higher potency of fresh ZnO NPs in inducing cell apoptosis.

Model for Fresh ZnO NPs but not aged ZnO NPs induces Caspase 8- and Caspase 3-dependent apoptosis. The increased expression of apoptotic gene CD69 activates Fas and apoptosis annexin V expression in fresh ZnO NP-exposed mammalian cells. The increased expression of apoptotic gene IL1α and IL1β partially activates Caspase 8-dependent apoptosis. It further causes activation of Caspase 3 and induces apoptosis. All these changes in mRNA and protein level were not detectable in aged ZnO NPs-exposed mammalian cells
Conclusiones
In the present study, the natural physicochemical transformation of ZnO NPs in ultrapure water was confirmed, and variations in cytotoxicity induced by fresh &aged NPs were investigated. We focused on RNA sequencing data from our aged ZnO NP-treated A L cells and that of fresh NPs from database. We compared those signaling pathway specifically enriched in aged NP-treated group, which are different from that of fresh NP- or ZnCl2 -treated groups. Our data indicated that the lower cytotoxicity of aged ZnO NPs is closely related to its attenuated ability in inducing apoptosis, while the transcriptional regulation of the multiple pathways activated by NPs promotes the establishment of cellular homeostasis in mammalian cells.
Disponibilidad de datos y materiales
No aplica.
Abreviaturas
- NP:
-
Nanopowders
- ZnO:
-
Óxido de zinc
- Zn5 (CO3 ) 2 (OH)6 :
-
Hydrozincite
- Zn (OH)2 :
-
Zinc hydroxide
- ZnCl2 :
-
Zinc chloride
- ZnSO4 :
-
Sulfuro de zinc
- ZnHPO4 :
-
Zinc hydrogen phosphate
- Zn3 (PO4 ) 2 :
-
Zinc phosphate
- A L cells:
-
Human–hamster hybrid cells
- CHO cells:
-
Chinese hamster ovary cells
- Jurket cells:
-
Peripheral blood T lymphocyte cells
- HMDM cells:
-
Human monocyte-derived macrophages
- NIH-3T3cells:
-
Mouse embryonic cells
- MSTO cells:
-
Human lung cancer cells
- RPMI1640:
-
Roswell Park Memorial Institute 1640
- ICE:
-
Interleukin 1beta-converting enzyme
- CED-3:
-
Caenorhabditis elegans death gene
- IL1α:
-
Interleukin 1alpha
- IL1β:
-
Interleukin 1beta
- mRNA:
-
Messenger ribonucleic acid
- cDNA:
-
Complementary deoxyribonucleic acid
- FBS:
-
Suero fetal bovino
- TEM:
-
Microscopía electrónica de transmisión
- XRD:
-
Difracción de rayos X
- RT-PCR:
-
Real-time polymerase chain reaction
- CPI:
-
The Nanotechnology Consumer Product Inventory
- XANES:
-
Synchrotron radiation X-ray absorption near-edge structure spectroscopy
- RIPA:
-
Radio immunoprecipitation assay
- SDS-PAGE:
-
Polyacrylamide gel electrophoresis
- PVDF:
-
Fluoruro de polivinilideno
- ECL:
-
Enhanced chemiluminescence
- CCK-8:
-
Cell counting kit-8
Nanomateriales
- Nanopartículas de FePO4 biocompatibles:administración de fármacos, estabilización de ARN y actividad funcional
- Efectos del espesor de la bicapa en las propiedades morfológicas, ópticas y eléctricas de los nanolaminados de Al2O3 / ZnO
- Ajuste de las morfologías de la superficie y las propiedades de las películas de ZnO mediante el diseño de la capa interfacial
- Aptitud ecológica en la síntesis y estabilización de nanopartículas de cobre:actividades catalíticas, antibacterianas, citotóxicas y antioxidantes
- Fabricación y caracterización de nanoclips de ZnO mediante el proceso mediado por poliol
- Un escrutinio comparativo in vivo de nanopartículas de óxido de zinc y cobre biosintetizadas por vías de administración intraperitoneal e intravenosa en ratas
- Síntesis verde de nanopartículas de metal y óxido de metal y su efecto sobre el alga unicelular Chlamydomonas reinhardtii
- Propiedades de las nanopartículas de óxido de zinc y su actividad contra los microbios
- Un enfoque fácil para la preparación de óxido de zinc de tamaño nanométrico en agua / glicerol con fuentes de zinc extremadamente concentradas
- Preparación y propiedades antibiofilm de películas compuestas de óxido de zinc / alúmina anódica porosa
- El papel de la robótica y la automatización en la Industria 4.0