


Información sobre la absorción celular y el tráfico intracelular de nanopartículas
Resumen
La ciencia de las nanopartículas está cambiando rápidamente el panorama de varios campos científicos y definiendo nuevas plataformas tecnológicas. Esto es quizás incluso más evidente en el campo de la nanomedicina, en el que las nanopartículas se han utilizado como herramienta para el tratamiento y diagnóstico de muchas enfermedades. Sin embargo, a pesar del tremendo beneficio conferido, los inconvenientes comunes de esta tecnología son sus posibles efectos a corto y largo plazo en el cuerpo humano. Para comprender estos temas, se han realizado muchos estudios científicos. Esta revisión intenta arrojar luz sobre algunos de estos estudios y sus resultados. Los temas que se examinaron en esta revisión incluyen las diferentes vías de captación posibles de nanopartículas y las rutas de tráfico intracelular. Además, también se abordan el efecto de las propiedades fisicoquímicas de las nanopartículas, como el tamaño, la forma, la carga y la química de la superficie, en la determinación del mecanismo de absorción y la función biológica de las nanopartículas.
Introducción
Las nanopartículas (NP) son una subcategoría de nanomateriales que se encuentran actualmente a la vanguardia de la investigación de vanguardia en casi todos los campos imaginables debido a sus propiedades únicas y su tremenda aplicabilidad [1, 2, 3, 4]. En un informe de investigación de mercado de tecnología titulado “Global NP Market Outlook 2020” por RNCOS, se informó que el mercado de NP crecerá a una tasa de crecimiento anual compuesta (CAGR) del 16% durante 2015-2020. La tecnología NP ha encontrado un nicho único en el campo de la biomedicina y la biotecnología con su repertorio de aplicaciones en rápido crecimiento [5, 6]. Por ejemplo, las NP se han aplicado para la administración de fármacos y genes [7, 8], la biodetección de patógenos [9], la detección de proteínas [10], la ingeniería de tejidos [11, 12], la obtención de imágenes de tumores y la orientación [13], la destrucción de tumores a través de hipertermia [14] y realce de contraste de resonancia magnética [15].
Debido a su pequeño tamaño, las NP pueden entrar fácilmente en las células y trasladarse a través de las células, tejidos y órganos. Las NP se utilizan ampliamente en aplicaciones biomédicas porque pueden atravesar la barrera biológica y entrar en la célula para ejercer su función. Sin embargo, como un arma de doble filo, los riesgos potenciales (es decir, el efecto adverso) de NP también surgen de esta capacidad [16, 17]. A pesar de su tamaño “pequeño”, las NP como moléculas polares no pueden difundirse a través de la membrana celular (MC). Dado que la MC es principalmente permeable a moléculas pequeñas y no polares, las NP emplean vías endocitóticas para ingresar a las células [18, 19]. La forma en que las NP ingresan a la célula es un factor clave para determinar sus funciones biomédicas, biodistribución y toxicidad. En nanomedicina, la entrada segura de NP en las células es un paso crucial para obtener una alta eficacia terapéutica. Además, el tráfico intracelular y el destino de las NP es un proceso vital para el éxito de las NP, teniendo en cuenta que estos portadores tienen como objetivo apuntar a un compartimento subcelular específico y entregar biomoléculas específicas como agentes de contraste, genes y fármacos [18, 20,21,22 ]. Más importante aún, la inducción de citotoxicidad por NP está determinada por su vía de entrada y localización intracelular. Por lo tanto, comprender la captación celular y el tráfico intracelular de NP es crucial en el diseño de nanomedicinas seguras y eficientes [23].
La captación celular, la focalización y el tráfico intracelular de NP pueden optimizarse ajustando las propiedades fisicoquímicas de las NP, como el tamaño, la forma y las propiedades de la superficie [24]. Por tanto, el conocimiento de los mecanismos subyacentes implicados en la captación celular es fundamental para evaluar el destino de las NP y su toxicidad. Esta revisión destaca las diferentes vías de captación posibles de NP y sus rutas de tráfico intracelular. Además, también se aborda el efecto de las propiedades fisicoquímicas de NP, como el tamaño, la forma, la carga y la química de la superficie sobre su internalización por las células. Comprender las propiedades fisicoquímicas de las NP en relación con su mecanismo de captación celular nos permitirá diseñar NP funcionales que son cruciales en aplicaciones biomédicas, como la entrega de cargas útiles de fármacos en el sitio de acción objetivo de una manera controlada con efectos tóxicos mínimos en los tejidos sanos circundantes. y órganos.
Vías de captación celular de NP
La MC, también conocida como membrana plasmática, encierra el citoplasma separando el líquido intracelular del extracelular. La CM es inmensamente importante ya que protege los componentes intracelulares, mantiene la homeostasis celular, confiere soporte estructural y retiene la composición de la célula [25,26,27,28,29]. CM consta de fosfolípidos dispuestos en una bicapa con proteínas incrustadas. Estas bicapas de fosfolípidos, con sus cabezas hidrófilas y sus colas hidrófobas, permiten la entrada de pequeñas biomoléculas. Más específicamente, el CM es una barrera selectivamente permeable que controla el paso de sustancias al interior de la célula [30, 31]. El MC emplea diferentes mecanismos de intercambio de sustancias que se dividen principalmente en dos categorías:transporte pasivo y transporte activo. Los gases como el oxígeno y el dióxido de carbono, las moléculas hidrófobas como el benceno y las moléculas sin carga como el agua y el etanol se difunden a través de la membrana desde las regiones de mayor a menor concentración. Este tipo de transporte que se realiza a lo largo del gradiente de concentración y se produce sin asistencia de energía se denomina transporte pasivo. Por el contrario, el transporte activo se produce contra el gradiente de concentración mediante el uso de energía proporcionada por el trifosfato de adenosina (ATP) [32,33,34,35,36].
Las biomoléculas polares o cargadas que no pueden atravesar la membrana plasmática hidrófoba se internalizan mediante una forma de transporte activo que se denomina endocitosis. En este proceso, la célula envuelve los materiales dentro del líquido extracelular por invaginación de CM y brota dentro de la célula, formando una vesícula rodeada de membrana llamada endosoma [37]. La endocitosis se puede clasificar básicamente en dos categorías principales:fagocitosis y pinocitosis. La fagocitosis (ingestión de células) es el proceso de absorción de desechos, bacterias u otros solutos de gran tamaño por parte de células de mamíferos especializadas llamadas fagocitos (es decir, monocitos, macrófagos y neutrófilos) [38, 39].
Una parte integral de la fagocitosis es un proceso llamado opsonización por el cual las opsoninas, como las inmunoglobulinas y las proteínas del complemento, recubren los materiales diana para activar los fagocitos de su presencia e inicializar la actividad fagocitótica [40]. A medida que el fagocito comienza a ingerir el material diana, simultáneamente estimulará la formación de una vesícula unida a la membrana llamada fagosoma en la que los materiales ingeridos se compartimentan dentro del fagocito. En las últimas etapas de este proceso, el fagosoma se fusionará con el lisosoma y los materiales son digeridos a pH ácido por las enzimas hidrolíticas contenidas en la luz lisosomal [41,42,43].
En todos los tipos de células, las partículas pequeñas dentro del rango de nanómetros son internalizadas por pinocitosis [44]. En la pinocitosis, la membrana plasmática de "bebida celular" forma una invaginación para absorber una pequeña gota de líquido extracelular que incluye moléculas disueltas en él. La pinocitosis no es un proceso discriminatorio y ocurre en casi todas las células de manera continua independientemente de las necesidades de la célula. Las sustancias atrapadas se comprimen en pequeñas vesículas que se denominan pinosomas, que se fusionan con los lisosomas para hidrolizar o descomponer el contenido [45, 46]. La fagocitosis y la pinocitosis se pueden distinguir por el tamaño de sus vesículas endocitóticas; el primero abarca la captación de partículas grandes por vesículas grandes con un tamaño de 250 nm, y el segundo abarca la captación de fluidos a través de vesículas pequeñas con un tamaño en el rango de unos pocos nanómetros a cientos de nanómetros [42, 47]. La pinocitosis se puede subdividir en endocitosis mediada por clatrina, endocitosis mediada por caveolas, endocitosis y macropinocitosis independientes de clatrina y caveolas [48, 49].
La endocitosis mediada por clatrina es el mecanismo de entrada celular para internalizar moléculas específicas en las células. Esta ruta de entrada ayuda a las células a absorber los componentes de la membrana plasmática y los nutrientes, incluido el colesterol por el receptor de lipoproteínas de baja densidad y el hierro por el receptor de transferrina [50,51,52,53,54,55,56]. En este proceso, ligandos particulares en el líquido extracelular se unen a los receptores en la superficie del CM formando un complejo ligando-receptor. Este complejo ligando-receptor se mueve a una región especializada de la MC que es rica en clatrina, por lo que son engullidos mediante la formación de vesículas recubiertas de clatrina. Una vez dentro de la célula, los recubrimientos de clatrina en el exterior de las vesículas se expulsan antes de fusionarse con los primeros endosomas. La carga dentro de los endosomas tempranos llegará eventualmente a los lisosomas a través de la vía endo-lisosomal [40, 57,58,59,60]. Cada tipo de NP es internalizado por la célula a través de una vía de captación preferencial. Por ejemplo, NP compuestas de poli (ácido láctico-co-glicólico), D, L-polilactida y poli (etilenglicolco-lactida) y sílice (SiO 2 ) Los nanomateriales basados en la clatrina son internalizados por la vía endocitótica mediada por clatrina [61]. Las NP de lípidos sólidos a base de cumarina son internalizadas por las células a través de una vía no dependiente de la energía, ya que la estructura de estas NP es similar a la CM. Todas las NP basadas en lípidos utilizan la vía de endocitosis mediada por clatrina [62]. Las NP de oro recubiertas de herceptina ingresan a la célula a través de una endocitosis mediada por receptores a través del receptor ErbB2 de membrana [63].
La endocitosis mediada por caveolas es la ruta de entrada celular que involucra invaginaciones de membrana en forma de matraz llamadas caveolas (pequeñas cuevas). Las caveolas están presentes en células endoteliales, epiteliales, adipocitos, células musculares y de fibroblastos [64,65,66,67]. El tamaño de las caveolas suele oscilar entre 50 y 80 nm y están compuestas por la proteína de membrana caveolina-1, que les confiere una estructura en forma de matraz [68,69,70,71]. La endocitosis dependiente de caveolas participa en la señalización celular y la regulación de proteínas de membrana, lípidos y ácidos grasos [61, 64, 67]. Una vez que las caveolas se desprenden de la membrana plasmática, se fusionan con un compartimento celular llamado caveosomas que existe a pH neutro. Los caveosomas pueden evitar los lisosomas y, por lo tanto, proteger el contenido de la enzima hidrolítica y la degradación lisosomal. Por lo tanto, los patógenos, incluidos los virus y las bacterias, utilizan esta ruta de entrada para evitar la degradación. Dado que la carga internalizada en las células por un mecanismo dependiente de la caveolina no termina en el lisosoma, esta vía se emplea en nanomedicina [54, 72,73,74].
La endocitosis independiente de clatrina y caveolas se produce en células privadas de clatrina y caveolina. Esta vía es utilizada por las hormonas del crecimiento, el líquido extracelular, las proteínas ligadas al glicosilfosfatidilinositol (GPI) y la interleucina-2 para ingresar a las células. Por ejemplo, el ácido fólico que emplea una vía independiente de clatrina y caveolas para entrar en las células [58, 72, 75,76,77,78,79] se conjuga con NP y polímeros utilizados en sistemas de administración de fármacos y como agentes de formación de imágenes [53 , 80, 81]. La macropinocitosis es un tipo de mecanismo de pinocitosis en el que las células absorben grandes volúmenes de líquido extracelular formando una vesícula grande (0,5-10 µm) llamada macropinosomas [82,83,84,85]. La macropinocitosis es una vía para internalizar células apoptóticas y necróticas, bacterias y virus, así como la presentación de antígenos. Esta vía puede internalizar NP del tamaño de una micra que no es posible que sean introducidas en las células por la mayoría de las otras vías. La macropinocitosis puede ocurrir en casi cualquier célula excepto en las células endoteliales de microvasos cerebrales [86,87,88,89]. Las NP ingresan a la célula a través de una de estas rutas endocitóticas, como se muestra en la Fig. 1.
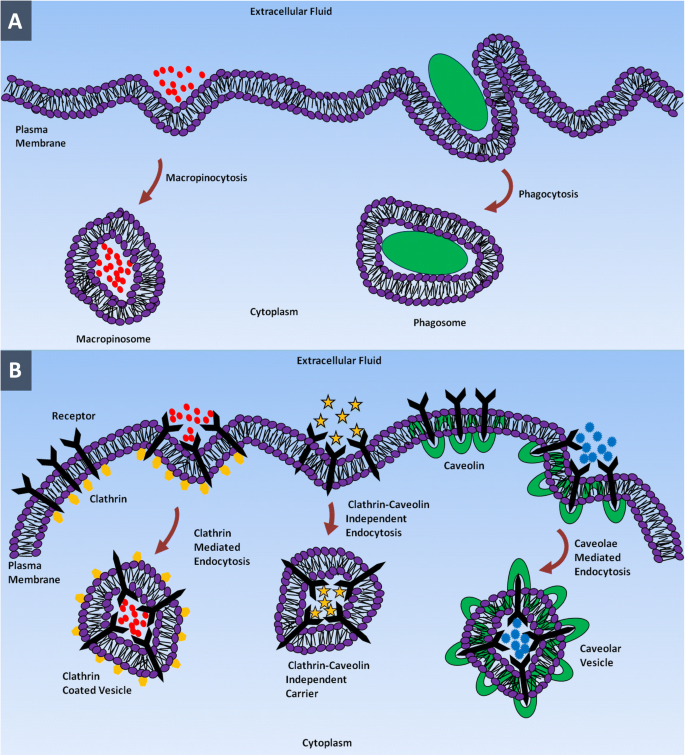
Entrada de NP en la célula mediante diferentes vías endocitóticas. un Macropinocitosis y fagocitosis. b Endocitosis mediada por clatrina, endocitosis independiente de clatrina-caveolina y endocitosis mediada por caveolas
Efecto de las propiedades fisicoquímicas de NP en la absorción celular
Estudiar el efecto de las propiedades fisicoquímicas de las NP como el tamaño, la forma, la carga superficial, la hidrofobicidad / hidrofilicidad de la superficie y la funcionalización de la superficie sobre la captación celular es crucial, ya que estos parámetros afectan directamente el nivel de captación, la ruta endocitótica y la citotoxicidad de las NP. [90, 91]. Los factores fisicoquímicos que afectan la captación celular de NP se ilustran en la Fig. 2. En la siguiente sección, se discute el impacto de estos parámetros en las interacciones célula-NP.
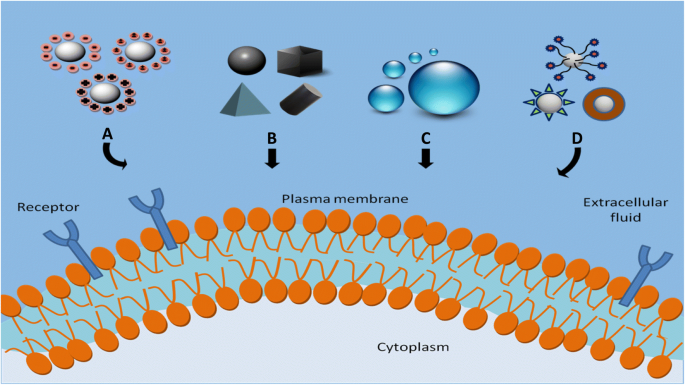
Factores fisicoquímicos que afectan la captación celular de NP. un Carga de superficie, b forma, c tamaño y d química de superficie
Efecto del tamaño
El tamaño de NP es un factor clave para determinar la eficiencia de la absorción celular [92], así como su potencial tóxico en las células vivas [24]. Además, se descubrió que el tamaño de NP también juega un papel importante en la determinación de la vía de captación. Las NP pequeñas con tamaños que van desde unos pocos a varios cientos de nanómetros ingresan a las células a través de pino o macropinocitosis. Se ha demostrado que las NP en el rango de tamaño de 250 nm a 3 μm tienen una fagocitosis in vitro óptima, mientras que las NP con el rango de tamaño de 120 a 150 nm se internalizan a través de endocitosis mediada por clatrina o caveolina, y el tamaño máximo de NP empleando esta vía se informó que era de 200 nm [47, 93]. En la vía mediada por caveolas, el tamaño de las caveolas dificulta la captación de NP más grandes [16, 17]. Un tipo particular de NP puede utilizar múltiples vías endocíticas según su tamaño.
Varios estudios han indicado que para la captación celular de NP, existe un tamaño óptimo de 50 nm en el que las NP se internalizan de manera más eficiente y tiene una tasa de captación más alta. Se demostró que la absorción de NP disminuye para partículas más pequeñas (alrededor de 15-30 nm) o partículas más grandes (alrededor de 70-240 nm) [94,95,96,97,98,99]. Además, las NP que oscilan entre 30 y 50 nm interaccionan eficazmente con los receptores de CM y, posteriormente, se internalizan mediante endocitosis mediada por receptores [97]. En la aplicación de administración de fármacos de NP, la principal preocupación es evitar que las NP sean eliminadas por el sistema reticuloendotelial y prolongar su tiempo de circulación en la sangre, mejorando así la biodisponibilidad en el objetivo. En este sentido, aumentar el tamaño de los NP conducirá a un aumento en la tasa de aclaramiento [100,101,102,103,104,105]. Por lo tanto, comprender el papel del tamaño de NP en la captación celular es crucial para diseñar NP efectivas y seguras para aplicaciones médicas.
Aunque diferentes estudios han investigado la relación entre el tamaño de NP y las vías de captación, los resultados revelados siempre han sido inconsistentes [93, 106,107,108,109]. Estas contradicciones pueden estar relacionadas con la complejidad de controlar otros parámetros de NP durante el proceso de control de tamaño. Además de eso, los tamaños de los NP medidos después de la síntesis pueden sufrir cambios durante los estudios in vitro e in vivo debido a la aglomeración y agregación que, a su vez, podrían afectar las vías de internalización celular [110, 111]. El impacto del tamaño de partícula en la vía de captación celular en células B16 no fagocíticas se investigó empleando diferentes tamaños de perlas de látex fluorescentes en el rango de 50-1000 nm [93]. Los resultados han demostrado que el mecanismo de internalización de estas perlas depende significativamente del tamaño de partícula. En particular, las perlas con tamaños de 200 nm o menos fueron absorbidas por hoyos recubiertos de clatrina, mientras que las perlas más grandes fueron internalizadas por endocitosis mediada por caveolas. Lai y colaboradores [16] han descubierto que las NP poliméricas pequeñas con tamaños inferiores a 25 nm emplean un nuevo mecanismo para alcanzar la región perinuclear de las células a través de una vesícula no degradante fuera de la vía endo / lisosomal. Esta vía no está mediada por clatrina ni caveolas y es independiente del colesterol.
Se demostró que la captación de NP de oro (Au) de diferentes tamaños (2 a 100 nm) conjugadas con Herceptin-AuNP por las células SK-BR-3 depende del tamaño. La internalización celular más alta se observó para las NP en los rangos de tamaño de 25 a 50 nm [63]. En esta ruta de entrada, se encontró que el tamaño de NP es el determinante en la unión y activación de los receptores de membrana y la expresión final de las proteínas. Se evaluó el efecto de la variación en el tamaño y la forma de las AuNP coloidales sobre la captación intracelular [112]. Se incubaron AuNP de tamaño de 14, 50 y 74 nm con forma esférica y de varilla con células HeLa. Se encontró que la absorción de NP depende en gran medida de su tamaño y forma y aquellas partículas con un tamaño de 50 nm mostraron la tasa de absorción más alta. Además, la absorción de AuNP esféricos fue 500% más que NP en forma de varilla de tamaño similar. Shan y col. [113] investigó la fuerza dependiente del tamaño de endocitosing AuNP con diámetros de 4, 12 y 17 nm por células HeLa. Los resultados revelaron que tanto los valores de fuerza de absorción como de desunión aumentan según el tamaño de los AuNP. La absorción de SiO 2 Se han estudiado NP de diferentes tamaños (50, 100 y 300 nm) por células A549 (células epiteliales pulmonares) mediante combinación de citometría de flujo, microscopía de fluorescencia y electrónica. Estos investigadores habían demostrado que la absorción de SiO 2 NP ha disminuido por tamaño [114].
Efecto de la forma
Además del tamaño, la forma del NP también juega un papel fundamental en la vía de captación, así como en el tráfico de NP. Chithrani y col. [112] estudiaron el efecto de la forma de las AuNP coloidales sobre la captación de células HeLa. El resultado reveló que las AuNP esféricas tenían una absorción cinco veces mayor que las AuNP en forma de varilla. En otro trabajo, los mismos investigadores investigaron el nivel de absorción de AuNP esférico y con forma de varilla recubierto de transferrina en tres líneas celulares diferentes; Células STO, células HeLa y células SNB19 [94]. Observaron que las AuNP esféricas eran internalizadas por todas las líneas celulares a una velocidad mayor que las AuNP en forma de bastón.
Con el fin de establecer el efecto de la forma in vivo, Geng et al. [115] emplearon filomicellas para evaluar las diferencias en el transporte y tráfico de filamentos flexibles con esferas en roedores. Los resultados revelaron que las filomicelias permanecían en la circulación unas diez veces más que las esféricas. Además, las filomicelias esféricas son internalizadas por las células más fácilmente que los filamentos más largos. Gratton et al. [106] demostraron el efecto de la forma de las partículas de hidrogel monodispersas sobre la captación en las células HeLa. Descubrieron que los NP con forma de varilla tenían las tasas de internalización más altas en comparación con las esferas, cilindros y cubos. En otro estudio, se investigó el impacto de la forma de las NP en la absorción celular mediante el empleo de NP de poliestireno (PS) en forma de disco, esféricas y en forma de varilla en las células Caco-2. El resultado demostró que los NP en forma de varilla y disco se internalizaron dos veces más que los NP esféricos. Llegaron a la conclusión de que la administración de fármacos mediada por NP puede avanzar si se considera la forma de las NP [116].
Xu y colaboradores [117] estudiaron el impacto de la forma en la absorción celular mediante la preparación de NP de hidróxido doble en capas (LDH) con isotiocianato de fluoresceína (FITC) en diferentes morfologías, como láminas hexagonales (50-150 nm de ancho lateral y 10-20 nm de espesor) y varillas (de 30 a 60 nm de ancho y de 100 a 200 nm de largo). Todas las morfologías se tomaron mediante endocitosis mediada por clatrina. La nanoesfera de LDH-FITC se retuvo en el citoplasma, mientras que las nanovarillas de LDH-FITC se movieron hacia el núcleo mediante microtúbulos. Dasgupta y col. aplicó [118] una simulación para probar el papel de la forma de las NP en la captación celular. Han simulado la envoltura de membranas de los NP en forma de nanobarra y nanocubo. Para las partículas en forma de bastón, encontraron estados endocitóticos estables con una fracción de envoltura pequeña y alta; el incremento en la relación de aspecto no era deseable para un envoltorio completo. Nangia y Sureshkumar [119] han informatizado el efecto de la forma en la tasa de translocación de los NP aplicando técnicas avanzadas de simulación de dinámica molecular. Una revelación importante del estudio es la variación significativa en la tasa de translocación de NP en forma de cono, cubo, varilla, arroz, pirámide y esfera.
Efecto de la carga superficial
Otro factor crítico que influye en la absorción celular de NP es la carga superficial. En la última década, se ha empleado la modificación de nano superficies para modificar la carga superficial de las NP para que sean catiónicas o aniónicas [92]. El CM cargado negativamente mejora la captación de NP cargadas positivamente. En particular, los NP cargados positivamente tienen mayor internalización que los NP neutrales y cargados negativamente [47, 120]. Sin embargo, la captación de NP con carga positiva puede alterar la integridad de la MC y conducir a un aumento de la toxicidad [121, 122]. En general, las NP con carga positiva inducen la muerte celular [123, 124]. Curiosamente, las NP con carga neutra reducirán la captación celular en comparación con las NP con carga negativa [110, 125, 126, 127]. Además, la internalización de NP con carga negativa conduce a la gelificación de las membranas, mientras que las NP con carga positiva provocan fluidez en la MC [128, 129]. Además de la tasa de absorción de NP, las cargas superficiales también afectan los mecanismos de absorción. Más específicamente, las NP con carga positiva son internalizadas principalmente por la célula a través de macropinocitosis, mientras que la endocitosis independiente de clatrina / caveolas es el mecanismo para la captación de NP con carga negativa [130]. Las vías de captación celular varían cuando la superficie de las AuNP está recubierta por moléculas orgánicas. Por ejemplo, las AuNP simples que están cargadas positivamente se internalizan mediante macropinocitosis y endocitosis mediada por clatrina y caveolina, mientras que las AuNP recubiertas con polietilenglicol (PEG) con carga negativa se internalizan principalmente a través de endocitosis mediada por caveolina y / o clatrina [131].
Li y Gu [132] estudiaron la interacción de NP neutrales y cargadas con CM mediante simulaciones de dinámica molecular. Se encontró que los NP cargados tenían una mejor adhesión a los CM en comparación con los NP neutrales. Además, al aumentar la densidad de carga de las NP, la membrana puede envolverlas completamente. Otro grupo de investigación empleó simulación de dinámica molecular para investigar las interacciones de AuNP catiónicas y aniónicas con CM. Los resultados han revelado que la alteración del CM debido a la penetración de AuNP aumenta a medida que aumenta la densidad de carga de los AuNP [133]. Estos hallazgos sugieren una forma de controlar las interacciones entre las células y AuNP mediante la manipulación de las densidades de carga superficial de las AuNP para optimizar su absorción y minimizar la citotoxicidad, que son características esenciales para cualquier NP que se esté considerando para aplicaciones biomédicas.
Li y Malmstadt [134] estudiaron la interacción de las PS-NP cargadas positiva y negativamente con la membrana biológica. El resultado mostró que la fuerte interacción electrostática entre NP catiónicas y los grupos fosfato de la membrana condujo a mejorar la unión NP-membrana y la tensión superficial de la membrana, lo que a su vez resulta en la formación de poros. Se informó que la tasa de absorción de AuNP cargados positivamente en células SK-BR-3 fue cinco veces mayor que la de AuNP cargados negativamente. Estos investigadores también han explorado que los AuNP cargados positivamente fueron internalizados por vías de no endocitosis, mientras que los AuNP cargados negativamente fueron absorbidos por las células a través de vías de endocitosis [135].
Hauck y col. [107] sondeó la absorción de nanobarras de oro (AuNR) con un rango de tamaño de 18 a 40 nm y cargas superficiales en el rango de + 37 mV a - 69 mV por las células HeLa. Los resultados indicaron que para todas las concentraciones de AuNR, la internalización más alta en las células HeLa fue con las cargas superficiales de + 37 mV y la internalización más baja a -69 mV. Huhn et al [136] evaluaron las interacciones dependientes de la carga de las AuNP coloidales con diferentes líneas celulares, como las células de fibroblastos 3T3, las células progenitoras neurales C17.2 murinas y las células endoteliales de la vena umbilical humana. El resultado mostró que para todas las líneas celulares, las AuNP catiónicas tenían una mayor absorción que la contraparte aniónica. Llegaron a la conclusión de que la captación celular depende en gran medida del signo de carga. Además, el estudio de citotoxicidad indicó que, como consecuencia de una mayor absorción de las NP cargadas positivamente, muestran una mayor toxicidad que las cargadas negativamente.
Efecto de la hidrofobicidad
La hidrofobicidad de las NP es un factor determinante en su interacción con la MC [92, 137]. Varios estudios demostraron el impacto de la hidrofobicidad de los NP en sus interacciones con el CM. Li y col. [138] estudiaron el efecto de la hidrofobicidad / hidrofilicidad de las NP en la interacción con la CM mediante el empleo de simulaciones de dinámica molecular. Los resultados han revelado que los NP hidrófobos crearon inclusión en el CM, mientras que se encontró que los NP hidrófilos se adsorben en el CM. En otra investigación, se aplicó un enfoque de simulación para investigar el efecto de la hidrofobicidad en la interacción NP-célula. Se observó que las NP hidrófilas estaban envueltas, mientras que las NP hidrófobas se incrustaban dentro del núcleo hidrófobo interno de las bicapas al penetrar directamente en la membrana [139].
Las interacciones de QDNP con membranas mixtas de lípidos / polímeros se evaluaron cambiando la superficie de hidrofobicidad de las NP. Se observó que los NP hidrófobos se han ubicado dentro de los dominios del polímero en una monocapa mixta de lípidos / polímeros de las membranas, mientras que los QDNP hidrófilos se adsorben en las monocapas y se extienden por todas partes, lo que indica un mayor efecto sobre el empaquetamiento de moléculas en la interfaz aire / agua [140]. . Se estudió la incorporación de AuNP funcionalizados con ligandos hidrófobos e hidrófilos mixtos en las paredes de los liposomas. El resultado demostró que los ligandos hidrofóbicos interactúan con el núcleo hidrofóbico de la bicapa, mientras que los ligandos hidrofílicos interactúan con la solución acuosa [141].
Efecto de la modificación de la superficie
En aplicaciones biomédicas de NP, la modificación química superficial de NP es un paso crítico utilizado para disminuir la toxicidad, aumentar la estabilidad y para controlar y modular la internalización celular de NP, de ahí su destino biológico [142]. La funcionalización superficial de las NP comprende predominantemente PEG, el grupo carboxilo negativo (–COOH), grupos funcionales neutrales como los grupos hidroxilo (–OH) y el grupo amina positivo (–NH2). El incremento en la cantidad de (–NH2) conduce a una carga superficial positiva mejorada y, por lo tanto, aumenta la captación de NP en las células [143,144,145,146]. De manera similar, los grupos funcionales –COOH aumentan la carga negativa de las NP y, en consecuencia, mejoran su absorción [144].
Tao y col. [147] han diseñado un bioconjugado NP-aptámero funcionalizado con polidopamina para el direccionamiento tumoral. Han informado que los NP funcionalizados tienen una mejor eficacia de direccionamiento en comparación con los NP no funcionalizados, lo que indica tasas de captación celular más altas para los NP funcionalizados, lo que se traduce en un efecto terapéutico mejorado. En otra investigación, los NP funcionalizados con ácido fólico demostraron una mayor eficacia en la selección de células cancerosas de cuello uterino que los NP no funcionalizados [148]. Qiu y colaboradores [90] estudiaron el impacto del recubrimiento de la superficie sobre la toxicidad y la absorción celular de AuNP. Han revelado que el recubrimiento de la superficie es un factor clave para determinar la tasa de absorción celular, ya que los AuNR recubiertos con poli (cloruro de dialildimetilamonio) mostraron una mayor eficiencia en la internalización por parte de las células.
Jiang et al. [149] investigaron las diferencias en la absorción celular de poliestireno prístino (PS-NP) y NP de poliestireno funcionalizado con amino [149]. Los resultados han demostrado que las NP de poliestireno funcionalizadas con amino tienen una tasa de absorción más alta que las PS-NP, y las primeras se internalizaron principalmente a través de la vía mediada por clatrina y las últimas mediante endocitosis independiente de clatrina. Esta notable diferencia destaca el papel clave de la modificación química de la superficie en las interacciones celulares con las NP. Fullereno de superficie modificada, C 60 ( C ( COOH ) 2 ) 2 Los NP fueron internalizados por las células predominantemente a través de endocitosis de una manera dependiente del tiempo, la temperatura y la energía. Se descubrió que la endocitosis mediada por clatrina es la vía preferida para la internalización de C 60 ( C ( COOH ) 2 ) 2 NP [150].
Efecto de la elasticidad
La elasticidad de los juegos NP es un factor intrínseco que influye en su internalización por parte de las células. La elasticidad de los NP puede explicarse por su resistencia a los cambios cuando se le aplican fuerzas. Rigidez, dureza y rigidez son algunos de los términos que son sinónimos para describir la elasticidad de los NP. Un índice de medición que se utiliza para medir la elasticidad de los NP es el módulo de Young y la unidad de medida es Pascal (Pa). Según esta medición, un valor de módulo de Young más alto denota una elasticidad NP más alta y viceversa. Ejemplos de dispositivos o instrumentos analíticos que se utilizan para medir este valor en NP son el microscopio de fuerza atómica, el reómetro y el nanoindentador. Los NP que tienen valores elásticos más altos se denominan NP duros y ejemplos de estos son NP de oro, puntos cuánticos y NP magnéticos. Las NP que tienen valores elásticos más bajos se denominan NP blandas y ejemplos de ellas son hidrogeles, liposomas y polímeros biodegradables.
Numerosos estudios que se han centrado en este parámetro de NP con respecto a la captación celular han informado sobre la preferencia de las células para internalizar NP más rígidas de manera más eficiente en comparación con NP más blandas [151, 152]. Evidentemente, esta observación se atribuye a un menor gasto energético total de las membranas para envolver NP más rígidas en comparación con NP más blandas, aunque la energía de deformación requerida para envolver las NP varía a lo largo del proceso de internalización. Además, el modelado computacional de envoltura de membrana de NP con elasticidad variable realizado mediante simulación de dinámica molecular de grano grueso (CGMD) coincide con la observación experimental sobre los cambios de energía de deformación implicados en la internalización de NP rígidas y blandas [153]. However, there are also other studies that have reported on softer NPs being internalized more efficiently than stiffer NPs [154, 155] and intermediate elastic NPs internalized more efficiently compared to either stiff or soft NPs [156]. Hence, tuning the elasticity of NPs for better cellular internalization could be a valuable tool in biomedical applications such as drug delivery. A potential application was demonstrated by Guo and coworkers, whereby accumulation of nanolipogels in tumour cells were enhanced primarily by controlling this parameter of NP [157].
Intracellular Trafficking of NPs
In the previous sections, different possible uptake pathways of NPs and the parameters that affect the efficacy of uptake has been discussed. Following uptake, the next crucial matter is the intracellular trafficking of NPs which determines its final destination within cellular compartments, its cytotoxicity and its therapeutic efficacy [158, 159]. After NPs are internalized by the cells, they will first encounter membrane-bound intracellular vesicles called early endosomes. Endosomes formed at the plasma membrane are categorized into three types; early endosomes, late endosomes and recycling endosomes [106, 160,161,162,163].
Early endosome ferries the cargo to the desired cellular destination. Part of the cargo is recycled to the plasma membrane via recycling endosomes. Early endosomes transform into late endosomes via maturation and differentiation process. The late endosomes will then integrate with lysosomes to form endolysosomal vesicles and hydrolytic enzymes contained within these vesicles degrade the trapped NPs [18, 164,165,166]. However, some NPs are able to escape this pathway and are released into the cytoplasm therefore bypassing the lysosomal degradation process [167,168,169]. Another intracellular degradation pathway which plays important role in the intracellular fate of NPs is an intracellular process called autophagy [170,171,172]. In this process, cytoplasmic contents will be surrounded by autophagosome and delivered to the lysosome to be broken down and recycled [173]. In addition, aggregated proteins and dysfunctional organelles are degraded by autophagy to maintain cellular homeostasis. It is necessary to consider this pathway since recent studies demonstrated that several NPs are capable of inducing autophagy [174,175,176,177,178].
The intracellular trafficking of Tat peptide-conjugated quantum dots (Tat-QDs) in live cells was studied by Ruan and co-workers [179]. Dynamic confocal imaging showed that Tat-QDs interacted with negatively charged CMs leading to its internalization by macropinocytosis. The QD containing vesicles were observed to be actively transported by molecular motors towards the perinuclear region known as the microtubule-organizing center (MTOC). Tat-QDs bind to cellular membrane structures such as filopodia and vesicle shedding results in releasing QD-containing vesicles from the tips of filopodia.
The uptake and intracellular fate of fluorescent carboxylated polystyrene particles (20 nm and 200 nm in diameter) were evaluated by applying it on hepatocyte [180]. It was found that the particles were internalized by hepatocytes in size, time and serum-dependent manner. The fate of the particles was studied and they were not observed in early endosomes or lysosomes, but only in the mitochondria of the hepatocyte. Particles accumulated inside bile canaliculi show that NPs can be eliminated within bile. A study on the uptake and intracellular fate of silver NPs into human mesenchymal stem cells demonstrated that they agglomerate in the perinuclear region [181]. It was observed by using fluorescent probes that particles are contained within endo-lysosomal structures but not in the cell nucleus, endoplasmic reticulum or Golgi complex. Confocal imaging of FITC conjugated titania nanotubes in mouse neural stem cells revealed that they have crossed the karyotheca entering the cell nucleus [182]. Single-walled carbon nanotubes were observed to enter the cytoplasm and localize in the cell nucleus leading to cell mortality [183]. Translocation of AuNRs towards the nucleus has also been reported [184].
Conclusiones
The application of NPs in the modern world is growing at an exponential rate as the scientific enterprise is looking for novel ways to address current problems. NPs can be found as active ingredients in many formulations intended for human consumption, from cosmetics to processed foods. As its application increases in consumer products, so does human exposure to NPs. Hence, more research should be carried out to understand its potential hazards to humans and other living beings. In this review, we have looked at the current knowledge on the effects of NPs at a cellular level. Some of the topics discussed include cellular pathways of NPs and the influences of physiochemical properties of NPs on the uptake rate and uptake mechanism.
Abreviaturas
- ATP:
-
Adenosine triphosphate
- CAGR:
-
Compound annual growth rate
- CM:
-
Cell membrane
- FITC:
-
Fluorescein isothiocyanate
- GPI:
-
Glycosylphosphatidylinositol
- LDH:
-
Layered double hydroxide
- MTOC:
-
Microtubule-organizing center
- NP:
-
Nanoparticle
- PEG:
-
Polyethylene glycol
Nanomateriales
- Nanopartículas para la terapia del cáncer:avances y desafíos actuales
- Nanopartículas de FePO4 biocompatibles:administración de fármacos, estabilización de ARN y actividad funcional
- Preparación y propiedades magnéticas de nanopartículas de espinela de FeMn2O4 dopadas con cobalto
- La preparación de la nanoestructura yema-cáscara de Au @ TiO2 y sus aplicaciones para la degradación y detección del azul de metileno
- Poliglicerol hiperramificado modificado como dispersante para el control del tamaño y la estabilización de nanopartículas de oro en hidrocarburos
- Síntesis y rendimiento in vitro de nanopartículas de hierro-platino recubiertas de polipirrol para terapia fototérmica e imágenes fotoacústicas
- Un escrutinio comparativo in vivo de nanopartículas de óxido de zinc y cobre biosintetizadas por vías de administración intraperitoneal e intravenosa en ratas
- Preparación de NP de Ag encapsulado de silicalita-1 poroso jerárquico y su rendimiento catalítico para la reducción de 4-nitrofenol
- Síntesis verde de nanopartículas de metal y óxido de metal y su efecto sobre el alga unicelular Chlamydomonas reinhardtii
- Sobreenfriamiento de agua controlado por nanopartículas y ultrasonido
- Potencial de activación antiproliferativa y de apoptosis de nanopartículas de lípidos dirigidos a base de paclitaxel con internalización celular mejorada por receptores de transferrina:un estudio …