


Roles de ROS y detención del ciclo celular en la genotoxicidad inducida por nanoestructura de núcleo de nanobarra de oro / capa de plata
Resumen
Para comprender la genotoxicidad inducida en el hígado por las nanopartículas de plata (AgNP) y los iones de plata, se utilizaron en este estudio una nanoestructura de núcleo de nanovarilla de oro / capa de plata (Au @ Ag NR) y hepatocitos humanizados HepaRG. La participación del estrés oxidativo y la detención del ciclo celular en el ADN y el daño cromosómico inducido por 0.4-20 µg mL −1 Au @ Ag NR se investigaron mediante ensayo cometa, ensayo γ-H2AX y ensayo de micronúcleos. Además, se analizó la distribución de Au @ Ag NR. Nuestros resultados demostraron que tanto Ag + y Au @ Ag NR condujeron a la escisión del ADN y daño cromosómico (clastogenicidad) en las células HepaRG y que el Au @ Ag NR retenido en el núcleo puede liberar más Ag + , agravando los daños, que son causados principalmente por la detención del ciclo celular y la formación de ROS. Los resultados revelan la correlación entre la acumulación intracelular, Ag + liberación de iones y la potencial genotoxicidad de los AgNP.
Introducción
Las nanopartículas de plata (AgNP), de 1 a 100 nm de tamaño, pueden exhibir un amplio espectro de propiedades antimicrobianas al penetrar patógenos e inactivar el grupo sulfhidrilo interno de sus enzimas metabólicas [1]. Han demostrado una potente bacteriostasis y efectos bactericidas en Escherichia Coli , Neisseria gonorrhoeae y Chlamydia trachomatis , y se emplean ampliamente como recubrimientos médicos, productos domésticos [2] y apósitos para heridas [3]. Evidencia convincente muestra que las nanopartículas son capaces de entrar en el núcleo e interferir con el proceso de síntesis y transcripción del ADN [4]. En nuestro estudio anterior, informamos que una sola dosis intravenosa de 5 mg / kg de AgNPs podría introducir una rotura cromosómica notable en las células de la médula ósea de ratas Sprague-Dawley [5]. Una sola inyección intraperitoneal de 10 mg / kg o más de AgNP indujo daños tanto en el ADN como en los cromosomas [6]. Flower y col. [7] sugirió que los AgNP en dosis de 50 y 100 μg mL −1 podría desencadenar daños en el ADN dentro de los cinco minutos posteriores a la administración, lo que resalta la genotoxicidad de la plata (Ag) de liberación rápida. Teniendo en cuenta el riesgo de exposición excesiva, la investigación de la nanogenotoxicología o el daño del ADN y el potencial carcinogénico de los nanomateriales artificiales ha recibido mucha atención [8].
Se considera que los principales mecanismos de las lesiones genéticas inducidas por AgNP son la sobreproducción de especies oxidativas reactivas, la inflamación y la alteración del ciclo celular [9, 10]. Como se sugirió en estudios anteriores, los AgNP podrían interactuar directamente con el ADN a través del daño oxidativo [11] e interferir en la interfase a nivel del ADN y la mitosis a nivel cromosómico, o interactuar con la nucleoproteína y el aparato del huso mitótico para perturbar los puntos de control del ciclo celular [ 12]. Sin embargo, si la genotoxicidad inducida por los AgNP es parcialmente atribuible a las nanopartículas [13, 14] o completamente al Ag + liberado iones aún no está claro [15, 16].
La investigación de la genotoxicidad de los AgNP es difícil debido a la liberación inestable e ininterrumpida de la plata en los tejidos, lo que dificulta la localización de los AgNP y la diferenciación del nanocore del Ag. Nuestro grupo ha desarrollado recientemente una nanoestructura de núcleo de nanobarra de oro / capa de plata (Au @ Ag NR) para estudiar la toxicidad inducida por nanopartículas [17]. El núcleo de oro de Au @ Ag NR es fisiológicamente innato en el tejido y podría usarse como un estándar interno para monitorear la liberación de Ag + iones de la barra mediante el seguimiento del cambio en la relación Ag / Au, medido mediante espectrometría de masas de plasma acoplado inductivamente (ICP-MS) [18]. Mediante este método, se pueden identificar los diferentes orígenes de las toxicidades. Estudios anteriores han demostrado que el Ag + publicado Los iones de la capa de Au @ Ag NR produjeron daño oxidativo renal y eventualmente condujeron a cambios morfológicos y deterioro de la función de filtración del glomérulo [19]. Jiang y col. [20] sugirió que tanto la actividad específica de partículas como la liberación de iones de plata intracelular por Au @ Ag NR contribuyen a la respuesta tóxica de las células de la granulosa. También adoptamos Au @ Ag NR como modelo para estudiar el potencial de genotoxicidad in vivo de los AgNP y demostramos que la clastogenicidad, y no la mutagenicidad, es la forma principal de genotoxicidad inducida por la capa de Ag y el Ag + iones, mientras que no hubo diferencias en sus patrones de toxicidad [21].
El hígado es uno de los principales órganos propensos a la acumulación de AgNP y es reconocido como un órgano / tejido diana de la genotoxicidad inducida por AgNP. Nuestro estudio anterior mostró que cierta cantidad de plata (8,26 ± 3,90 μg / g) y oro (80,07 ± 64,72 μg / g) permanecía en el hígado de ratas SD ocho semanas después de la administración intravenosa de una dosis de Au @ Ag NR [21 ]. En este estudio, intentamos identificar las funciones de la detención del ciclo celular y el estrés oxidativo reactivo en los daños cromosómicos y del ADN inducidos por AgNP utilizando Au @ Ag NR en células HepaRG derivadas de hepatoma humano. Los ensayos de genotoxicidad, incluido el ensayo cometa, el ensayo γ-H2AX y la prueba de micronúcleos, se realizaron en paralelo con el eliminador de radicales oxidativos para investigar la contribución de las especies reactivas de oxígeno (ROS) en el daño del ADN / cromosoma, mientras que la apoptosis celular, el ciclo celular y proteínas relacionadas se determinaron para explorar los mecanismos por los cuales los AgNPs interrumpen la síntesis y replicación del ADN. Además, se investigó la acumulación y distribución intracelular de Au @ Ag NR combinando espectrometría de masas de plasma acoplado inductivamente (ICP-MS) y microscopía electrónica de transmisión (TEM) para diferenciar el papel de las nanopartículas y los iones de Ag liberados.
Materiales y métodos
Cultivo y tratamiento celular
En este estudio se utilizó la línea celular de hepatoma humano HepaRG (Thermo Fisher Scientific). Las células se cultivaron en RPMI 1640 que contenía suero bovino fetal al 10% (FBS, Australia Origin, Gibco) y solución de penicilina-estreptomicina-glutamina al 1% (Gibco) en una atmósfera humidificada de CO 2 al 5% a 37 ° C. Las células se trataron con concentraciones crecientes de Au @ Ag NR durante 24 ho 72 h, respectivamente, y las concentraciones se determinaron de acuerdo con IC 50 estimado por ensayo de viabilidad celular. Para investigar el papel de ROS en la genotoxicidad, 1 mM N -Se aplicó acetil-l-cisteína (NAC, Sigma-Aldrich) durante 1 h antes del tratamiento con Au @ Ag NR.
Ensayo de viabilidad / crecimiento de células de ATP
Las células se sembraron en una placa de 96 pocillos a una densidad de 5 × 10 3 /bien. Después de 24 h de incubación, se aspiró el medio y las células se expusieron a diferentes concentraciones de Au @ Ag NR durante 24 ho 48 h, respectivamente. Se preparó un amplio espectro de concentraciones y se realizaron cuatro pozos por tratamiento en un período de tratamiento. La citotoxicidad de Au @ Ag NR se examinó mediante el ensayo de trifosfato de adenosina (ATP) (ensayo CellTiter-Glo® 2.0, Promega), que mide la actividad metabólica celular cuantificando la cantidad de ATP, un parámetro importante del metabolismo en células viables. Las señales luminiscentes, que reflejan la cantidad de células viables, se detectaron mediante el lector de placas VICTOR Multilabel Plate Reader (2030-0050, PerkinElmer) e IC 50 Los valores se estimaron como la concentración de Au @ Ag NR para la viabilidad media máxima mediante Prism 7 (GraphPad Prism 7, CA, EE. UU.). El índice de viabilidad se calcula utilizando la siguiente ecuación:
$$ {\ text {Viability}} \, {\ text {Ratio}} \, \ left (\% \ right) ={\ text {RLU}} _ {{{\ text {sample}}}} / { \ text {RLU}} _ {{{\ text {vehículo}}}} \ veces {1} 00 \% $$donde RLU es la unidad de luz relativa representada como el valor medio de cuatro pozos, RLU vehículo representaron células no tratadas con nanovarillas y RLU muestra representaron células que fueron tratadas con diferentes concentraciones de Au @ Ag NR.
Determinación de la concentración de plata y oro en celdas
Las muestras de células se digirieron en ácido nítrico utilizando el sistema de digestión por microondas. Tras la digestión, las muestras se prepararon con una mezcla que contenía ácido nítrico al 1% y ácido clorhídrico. Las cantidades de Ag y Au en las soluciones se determinaron mediante ICP-MS (NexION300X, PerkinElmer). Se utilizó el análisis TEM para determinar la presencia de Au NR y Au @ Ag NR en la célula. Las muestras de células se fijaron en una mezcla de glutaraldehído al 2,5% y paraformaldehído al 2% durante 2 ha 4 ° C. Los sedimentos celulares se fijaron y enjuagaron tres veces en tampón fosfato (pH 7,4) y se fijaron posteriormente en tetróxido de osmio al 1% durante 2 ha 4ºC. Posteriormente, las muestras se enjuagaron en agua destilada tres veces y se deshidrataron durante 15 min en diferentes concentraciones de etanol (50%, 70%, 90% y 100% de etanol, respectivamente) una tras otra. Posteriormente, se aplicó óxido de propileno en diluciones 1:1 y 1:3 a la resina a 20–26 ° C durante 2 h. La polimerización se realizó mediante calentamiento gradual a 35 ° C durante 16 h, 45 ° C durante 8 h, 55 ° C durante 14 hy 65 ° C durante 48 h. Las secciones ultrafinas se tiñeron durante 25 min con acetato de uranilo y citrato de plomo y se analizaron con un microscopio electrónico de transmisión (H-7650, HITACHI, Japón).
Ensayo cometa convencional y modificado
Las células se sembraron en placas de 12 pocillos a densidades de 2 × 10 5 / pozo o 3 × 10 5 / pocillo para un tratamiento de 24 o 72 h, respectivamente. Peróxido de hidrógeno (H 2 O 2 ) a una concentración de 200 µmol se expuso a las células como control positivo durante una hora. Para cada muestra, se prepararon dos pocillos tanto para el tratamiento convencional como para el tratamiento con formamidopirimidina glicosilasa (Fpg). El ensayo cometa convencional se realizó en condiciones alcalinas (pH> 13) como se describió anteriormente [21]. Para los pocillos tratados con Fpg, se aplicó un tratamiento adicional con Fpg antes del procedimiento de desenrollado del ADN y los portaobjetos se sumergieron en un tampón enzimático (KCl 0,1 M, EDTA 0,5 mM, HEPES 40 mM, 0,2 mg.mL -1 BSA) tres veces durante 5 minutos cada una. El Fpg (New England Biolabs, Inc., Reino Unido) se diluyó a 1:50.000 con tampón enzimático. Se agregaron alícuotas de cien mililitros de la enzima diluida a cada gel en los portaobjetos de microscopio y se incubaron en una cámara de humedad a 37 ° C durante 30 min. Los pasos restantes fueron los mismos que los del tratamiento convencional. Los ensayos del cometa se realizaron por triplicado. Al menos 50 células por muestra se puntuaron de forma independiente utilizando el microscopio fluorescente Nikon Eclipse 80i (Nikon, Tokio, Japón), mientras que se utilizó Komet 6.0 (Andor Technology, Belfast, Reino Unido) para analizar el valor medio del porcentaje de ADN en la cola y la cola de oliva. momento (OTM) de cada muestra.
Calificación de focos de γ-H2AX mediante citometría de flujo y detección de alto contenido
Para la cuantificación mediante citometría de flujo, las células se sembraron en placas de 12 pocillos a densidades de 2 × 10 5 / pozo o 3 × 10 5 / pocillo para un tratamiento de 24 o 72 h, respectivamente, mientras que para el ensayo de detección de alto contenido, las células se sembraron en placas de 96 pocillos a densidades de 6 × 10 3 / pozo o 1 × 10 4 / pocillo para un tratamiento de 24 o 72 h, respectivamente. Como control positivo, se aplicó metanosulfonato de metilo 2 µM (MMS, Sigma-Aldrich) en paralelo con las células durante una hora. Las células se enjuagaron en solución salina tamponada con tris (TBS) y se fijaron con paraformaldehído al 4% durante 15 min a temperatura ambiente. Después de lavar con TBS, las células se incubaron con 50 μL de metanol helado durante 30 min a -20 ° C. Las células se aclararon adicionalmente en TBS tres veces y se aplicó el reactivo de bloqueo (TBS que contenía Triton X-100 al 0,3% y suero de cabra al 10%) durante 1 h. El anticuerpo primario (anti-fosfo-H2AX Ser139 de ratón, Millipore) se diluyó a 1:200 con reactivo de bloqueo y se incubó con las células durante la noche a 4ºC. A continuación, la placa se volvió a enjuagar con TBS tres veces, y posteriormente se añadió el anticuerpo secundario (Alexa Fluor 488 anti-ratón de cabra, Life Technologies), diluido con el reactivo de bloqueo en una proporción de 1:20. Las muestras se mantuvieron en la oscuridad a temperatura ambiente durante 1 h, y 2 μg mL −1 (20 μL / pocillo) Se añadió DAPI (Invitrogen) a cada pocillo. La fluorescencia se midió utilizando una citometría de flujo (FACSCalibur, BD Bioscience, NJ, EE. UU.) O un sistema de análisis de alto contenido (Operetta CLS, PerkinElmer). Para el ensayo de citometría de flujo, se analizaron los datos de al menos 10.000 células por grupo y los experimentos se realizaron por triplicado; para el análisis de alto contenido, se analizaron 20 campos visuales en cada pocillo y al menos cinco pozos en cada grupo.
Ensayo de citoma de micronúcleo de bloqueo de citocinesis (CBMN-cyt)
CBMN-cyt se realizó de acuerdo con el procedimiento descrito por Fenech et al. [22]. Las células se sembraron en placas de 12 pocillos a densidades de 2 x 10 5 / pozo o 3 × 10 5 / pocillo para un tratamiento de 24 o 72 h, respectivamente. 0,2 μg ml −1 Se expuso mitomicina C (MMC, Tokyo Chemical Industry Co., Ltd. Japón) a las células como control positivo durante 24 h. 3 μg mL −1 Se aplicó citocalasina B después de un tratamiento de 24 o 72 h para bloquear el proceso de citocinesis, y las células se recolectaron después de 40 h. Las muestras se tiñeron con Giemsa al 5% después de la hipotonicidad con 0.075 mol L −1 precalentado KCl y fijación con una mezcla 3:1 de metanol y ácido acético. Se prepararon pocillos por triplicado por grupo y se examinaron al menos 1000 células binucleadas por pocillo.
Medición de MDA, contenido total de GSH y SOD
Las células se cultivaron en placas de 12 pocillos a densidades de 5 x 10 5 / pozo o 3 × 10 5 / pocillo para un tratamiento de 24 o 72 h, respectivamente. Posteriormente, las células se recolectaron y se enjuagaron tres veces con solución salina tamponada con fosfato (PBS). Las cantidades de malondialdehído (MDA) en los homogeneizados de células se determinaron utilizando un método basado en ácido tiobarbitúrico (Instituto de Bioingeniería de Nanjing Jiancheng, Nanjing, China). Las cantidades de glutatión total (GSH) y superóxido dismutasa (SOD) se determinaron usando los kits de cuantificación de glutatión total y ensayo de SOD (Dojindo Molecular Technologies, Inc. Kumamoto, Japón), respectivamente. Las densidades ópticas (O.D) de cada pocillo se midieron con el lector de placas VICTOR Multilabel Plate Reader (2030-0050, PerkinElmer).
Análisis citométrico de flujo para el ciclo celular
Las células se cultivaron en placas de 6 pocillos a densidades de 1 x 10 6 / pozo o 5 × 10 5 / pocillo para un tratamiento de 24 o 72 h, respectivamente, y posteriormente se fijaron con etanol al 70% a 4 ° C durante la noche. Las muestras se enjuagaron con PBS tres veces y se tiñeron con tampón de tinción PI / Rnasa (BD Biosciences) durante 15 min a temperatura ambiente. Se determinaron las poblaciones de células bajo la fase G0 / G1, S y G2 / M entre 20.000 células empleando regiones con área FL2 frente a ancho FL2. El análisis se realizó mediante citometría de flujo (FACSCalibur, BD Bioscience, NJ, EE. UU.) Y FlowJo (BD Bioscience), y los experimentos se realizaron por triplicado.
Análisis citométrico de flujo de apoptosis celular
Las células se cultivaron en placas de 6 pocillos a densidades de 1 x 10 6 / pozo o 5 × 10 5 / pocillo para un tratamiento de 24 o 72 h, respectivamente. Posteriormente se enjuagaron dos veces con PBS y se diluyeron con 500 μL de tampón de unión 1x (FITC Annexin V Apoptosis Detection Kit I, BD Bioscience) para ajustar la suspensión a aproximadamente 1x10 6 células / ml, y posteriormente se mezcló una dilución de 100 µL con 5 µL de FITC Anexina V y 5 µL de PI. Las muestras se tiñeron a temperatura ambiente durante 15 min y se analizaron al menos 10,000 células para determinar la población celular en apoptosis temprana y tardía empleando regiones con FL1H versus FL2H usando citometría de flujo (FACSCalibur, BD Bioscience, NJ, EE. UU.) Y FlowJo (BD Bioscience). Los experimentos se realizaron por triplicado.
Análisis de Western blot
Las células se cultivaron en un 2 de 75 cm matraz a densidades de 1 × 10 7 / pozo y 6 × 10 6 / pocillo para un tratamiento de 24 y 72 h, respectivamente. Las células se lisaron con tampón de lisis RIPA que contenía inhibidor de proteasa (PMSF) y se determinó la concentración de proteínas usando un kit de cuantificación de proteínas BCA (Beyotime Biotechnology, China). Las concentraciones de las muestras se ajustaron usando tampón de lisis RIPA antes de la desnaturalización calentando a 95 ° C durante 3 min. Las muestras de proteína se separaron por electroforesis en geles de poliacrilamida SDS al 12% y se transfirieron a membranas de nitrocelulosa (Millipore). Las membranas se bloquearon con leche desnatada al 5% durante 30 min y se incubaron con p53 primario (SC-137174, Santa Cruz), p21 (SC-6246, Santa Cruz) y β-actina (sc-47778, Santa Cruz) y anticuerpos secundarios. IgG (H + L) -HRP de cabra anti-ratón (SE131, solabio), respectivamente. Los niveles de expresión de las proteínas diana en las muestras se visualizaron utilizando un método de quimioluminiscencia mejorada (ECL) y se analizaron con el sistema ImageJ (Institutos Nacionales de Salud).
Análisis estadísticos
Los datos se presentaron como la media ± SEM. Se utilizó el análisis de varianza unidireccional (ANOVA) para probar la significación estadística de las diferencias entre los grupos de control negativo y tratados, seguido de la prueba de comparación múltiple de Dunnett utilizando SPSS (versión 22, IBM, Armonk, NY, EE. UU.), Y se consideraron los datos estadísticamente significativo en P <0,05. Las figuras se prepararon utilizando GraphPad Prism 7 para Windows (GraphPad Software, La Jolla, CA, EE. UU.).
Resultados
Caracterización de Au NR y Au @ Ag NR
Se diseñaron, prepararon y caracterizaron nanobarras de oro (Au NR), núcleos de nanovarillas de oro y nanoestructuras de capa de plata (Au @ Ag NR) como se describió anteriormente [21]. Brevemente, los diámetros y longitudes medios son 15,0 ± 2,5 nm, 66,7 ± 2,5 nm para Au NR y 26,2 ± 3,0 nm, 72,7 ± 8,9 nm para Au @ Ag NR. El espesor de la capa de Ag es de aproximadamente 5 nm. Los potenciales zeta de los Au NR recubiertos con PDDAC y los Au @ Ag NR dispersos en agua fueron 37,7 ± 1,6 mV y 52,5 ± 1,4 mV, respectivamente. La relación en peso Ag / Au de Au @ Ag NR preparado se estimó en 2,3. Los resultados de la caracterización se muestran en la Fig. 1.
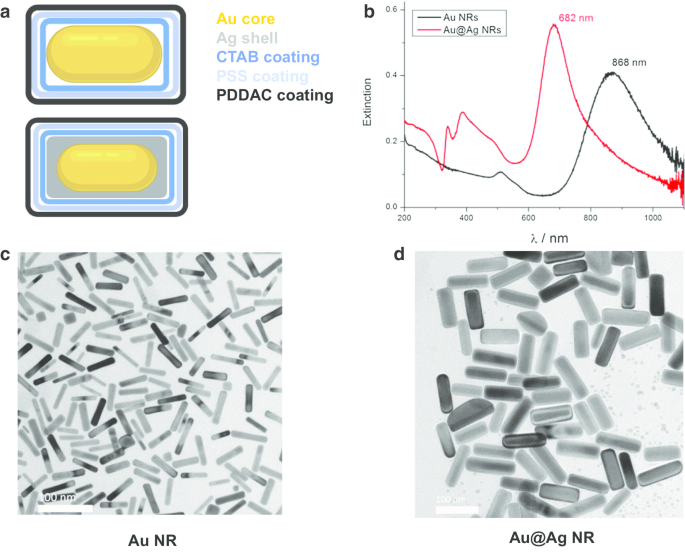
Caracterización de Au NR y Au @ Ag NR. un Diagrama estructural de Au NR y Au @ Ag NR; b Espectros de extinción UV-Vis-NIR de Au NR y Au @ Ag NR dispersos en agua; c imágenes TEM representativas de Au NR; d imágenes TEM representativas de Au @Ag NR
Viabilidad celular
La citotoxicidad de Au @ Ag NR hacia las células HepaRG se investigó mediante un ensayo de viabilidad de ATP (Tabla 1), y las células se expusieron a Au @ Ag NR durante 24 o 48 h en concentraciones que varían de 0,125 a 160 μg mL -1 . Au @ Ag NR indujo efectos citotóxicos significativos en formas dependientes del tiempo y de la dosis después de la exposición de 24 y 48 h, con un% de viabilidad IC 50 a 20 µg mL −1 y 6 µg mL −1 , instalado por el software GraphPad Prism 7.0, respectivamente. Considerando la citotoxicidad global, los períodos de tratamiento se ajustaron a 24 hy 72 h, mientras que las concentraciones aplicadas se determinaron en 0,8 µg mL −1 , 4 µg mL −1 y 20 µg mL −1 . Además, se incluyó Au NR como control inerte, y el contenido de Au en el grupo AuNR fue el mismo que 20 µg mL −1 Au @ Ag NR, que es 16 µg mL −1 . Por el contrario, el pretratamiento de NAC 1 mM se adoptó en el grupo Au @ Ag NR + NAC como control de la respuesta al estrés oxidativo (la concentración de Au @ Ag NR es de 20 µg mL −1 ).
Distribución celular de Au NR y Au @ Ag NR
La distribución del contenido de Au y Ag en las células HepaRG se analizó mediante ICP-MS. Como se muestra en las Tablas 2 y 3, el contenido de Ag aumentó de manera dependiente de la dosis. Sin embargo, el antioxidante N -La acetil-l-cisteína (NAC) como eliminador de radicales libres puede restringir la absorción celular de nanopartículas, ya que se observó un menor contenido de Ag a pesar de la misma concentración de Au @ Ag NR (20 µg mL −1 ) se aplicó en este grupo. La disminución en la relación Ag / Au de 24 a 72 h indicó una liberación continua de Ag + de la cáscara de Au @ Ag NR. Además, la captación celular de Ag es mucho mayor que la de Au (Tabla 4). Además, los datos de TEM mostraron que la mayoría de Au NR y Au @ Ag NR se retuvieron en las células como aglomerados. Las estructuras de nanobarras se vieron claramente dentro de las células sujetas a la exposición de Au NR o Au @ Ag NR sin entrar en el núcleo (Fig. 2).
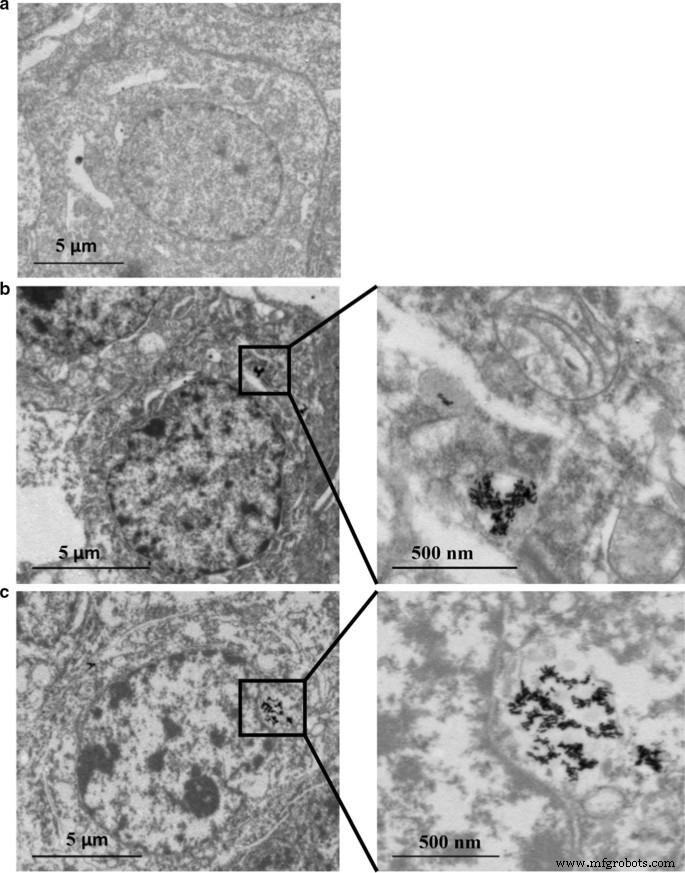
Internalización de Au NR y Au @ Ag NR:HepaRG por TEM a 80 kV después de 24 h de exposición a 16 μg mL −1 Au NR y 20 μg mL −1 Au @ Ag NR. un Control de vehículos; b Au NR; c Au @ Ag NR
Daño en el ADN
El daño al ADN desencadenado por Au @ Ag NR se evaluó mediante el ensayo cometa y el ensayo γH2AX (Fig. 3). Se observó en el ensayo cometa que de 0,8 a 20 µg mL −1 Au @ Ag NR podría introducir un daño significativo en el ADN. Después de una exposición de 24 o 72 h a Au @ Ag NR, tanto el% de ADN de la cola como el OTM de las células aumentaron de manera dependiente del tiempo y de la concentración. Además, se observó daño en el ADN asociado con la inducción de estrés oxidativo en las células tratadas con 20 µg mL −1 Au @ Ag NR por el ensayo cometa modificado con enzima Fgp (Fig. 3a, b). Para evaluar el grado de rotura de la doble cadena que representa una correlación más alta con la génesis del cáncer, se analizaron tanto las células positivas para γ-H2AX como las intensidades medias de fluorescencia en las células positivas para γ-H2AX. Después de una exposición de 24 h a Au @ Ag NR, no se encontraron diferencias entre los grupos de células positivas para γ-H2AX. Sin embargo, 4 μg mL −1 El grupo Au @ Ag NR provocó un aumento significativo después de un tratamiento de 72 h. Se observaron aumentos significativos en las intensidades de fluorescencia en todos los grupos de Au @ Ag NR después de 72 h en comparación con el control del vehículo (Fig. 3c-e, P <0.05).
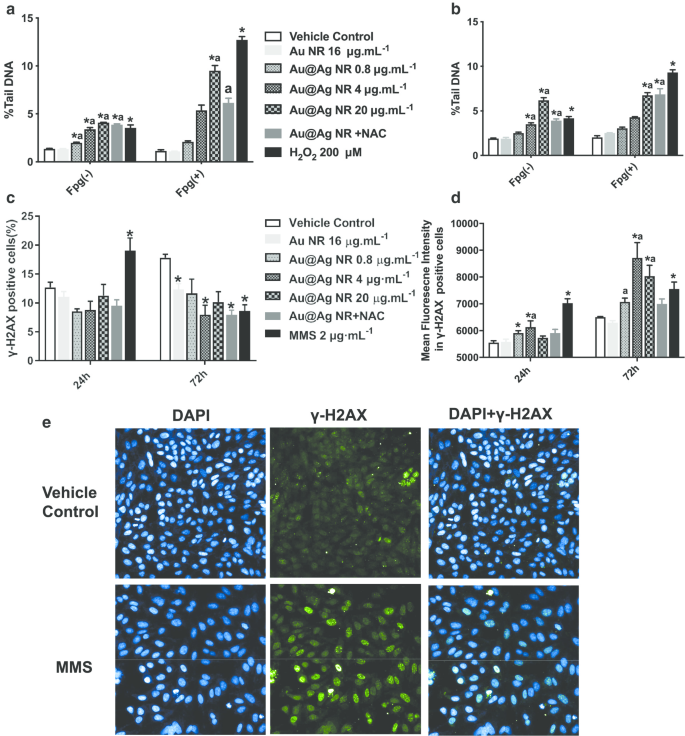
Daño del ADN inducido por Au @ Ag NR. Las células HepaGR se expusieron a Au @ Ag NR a diferentes concentraciones (0,8 a 20 μg mL −1 ) durante 24 hy 72 h, respectivamente. un % De ADN de cola promediado después de la exposición a Au @ Ag NR durante 24 h; b % de ADN de cola promediado después de la exposición a Au @ Ag NR durante 72 h; c porcentaje de células positivas con focos de γ-H2AX estimado mediante citometría de flujo; d intensidades medias de fluorescencia en células con focos de γ-H2AX estimadas mediante tinción inmunofluorescente. * P <0,05 frente al control del vehículo; a P <0,05 frente a Au NR. 2 μM mL −1 Se empleó MMS como control positivo
Daño cromosómico
La formación de micronúcleos es un biomarcador importante para identificar el daño cromosómico, que es un daño más crítico para el material genético que la rotura del ADN. La proporción de células binucleadas que contienen micronúcleos se puntuó como se muestra en la Fig. 4c. Au @ Ag NR aumentó la formación de micronúcleos en un patrón dependiente de la concentración. Después de una exposición de 24 h, las proporciones de micronúcleos observadas en las células tratadas con 4 μg mL −1 Au @ Ag NR y 20 μg mL −1 Au @ Ag NR fueron 1,133 ± 0,145% y 1,567 ± 0,318%, respectivamente, ambos significativamente más altos que los del grupo de control del vehículo. Después de una exposición de 72 h, la proporción de micronúcleos en las células tratadas con 4 μg mL −1 Au @ Ag NR fue 1,767 ± 0,233%, que fue significativamente mayor que el grupo de control de vehículo; la proporción de micronúcleos en las células tratadas con 20 μg mL −1 Au @ Ag NR fue 2.167 ± 0.252%, que fue significativamente más alto que los observados tanto en el grupo de control del vehículo como en 16 μg mL −1 Grupo Au NR (0,700 ± 0,153%). Por el contrario, no se encontraron diferencias entre las células tratadas con 20 μg mL −1 Au @ Ag NR + NAC y control del vehículo, lo que sugiere la participación de ROS en la rotura cromosómica inducida por Au @ Ag NR.
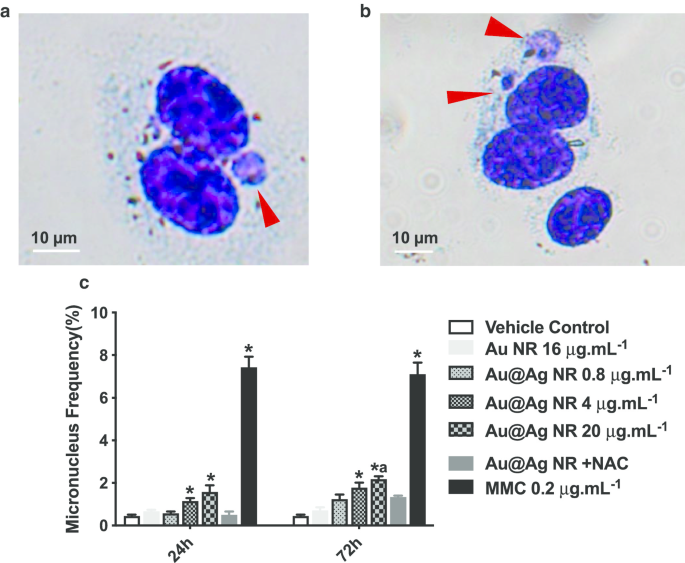
Daño cromosómico inducido por Au @ Ag NR. Las células HepaGR se expusieron a Au @ Ag NR a diferentes concentraciones de 0,8 μg mL −1 a 20 μg mL −1 durante 24 hy 72 h. un , b Imágenes representativas de micronúcleos (flecha roja); c frecuencia de micronúcleos (%). * P <0,05 frente al control del vehículo; a P <0,05 frente a Au NR. 0,2 μg ml −1 se utilizó mitomicina C como control positivo
Efectos de Au @ Ag NR en la formación ROS
Para explorar más a fondo el papel de la formación de ROS en el ADN inducido por Au @ Ag NR y los daños cromosómicos, se estimaron los niveles de MDA, GSH y SOD. Un aumento significativo en la formación de MDA ( P <0.05) después de la exposición a 20 μg mL −1 Au @ Ag NR durante 24 y 72 h (Fig. 5a). Además, los niveles de GSH y SOD en las células expuestas a Au @ Ag NR mostraron una reducción significativa ( P <0,05) de forma dependiente del tiempo y la concentración. Estos resultados sugirieron un desequilibrio entre la oxidación y la antioxidación, generado por la exposición de Au @ Ag NR (Fig. 5b, c).
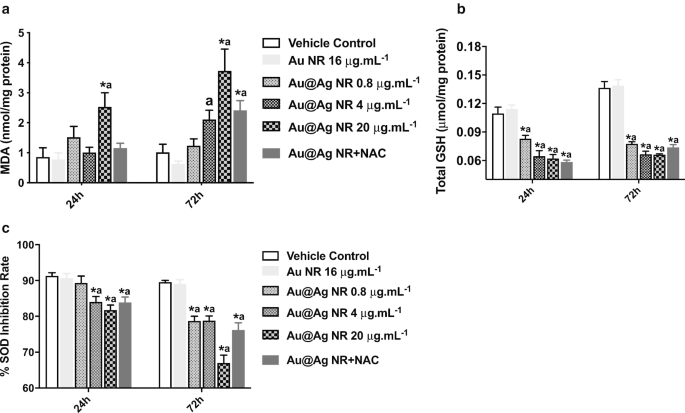
Efectos de Au @ Ag NR sobre la formación de ROS. Las células HepaGR se expusieron a Au @ Ag NR a diferentes concentraciones de 0,8 μg mL −1 a 20 μg mL −1 durante 24 hy 72 h. un Nivel de MDA; b Nivel de GSH; c Nivel de SOD. * P <0,05 frente al control del vehículo; a P <0,05 frente a Au NR
Efectos de Au @ Ag NR en el ciclo celular y la apoptosis
Después de una exposición de 72 h a Au @ Ag NR, se observó el aumento en el número de células en la fase G2 / M en 4 μg mL −1 Au @ Ag NR, 20 μg mL −1 Au @ Ag NR y Au @ Ag NR + NAC, con proporciones de 32,63% ± 1,77%, 32,267% ± 2,17% y 32,967% ± 4,25%, respectivamente (Fig.6a, b), que fueron significativamente mayores que las de el grupo de control del vehículo (22,37% ± 0,92%). Mientras tanto, se pudo observar la apoptosis celular inducida por Au @ Ag NR después de una exposición de 72 h, y la tasa de apoptosis tardía de las células tratadas con 20 μg mL −1 Au @ Ag NR y 20 μg mL −1 Au @ Ag NR + NAC fue 78,90 ± 1,19% y 70,20 ± 4,50%, respectivamente (Fig. 6c, d). Au @ Ag NR indujo más apoptosis tardía que apoptosis temprana, y el tratamiento de NAC podría aliviar la tasa celular de apoptosis tardía desencadenada por Au @ Ag NR.
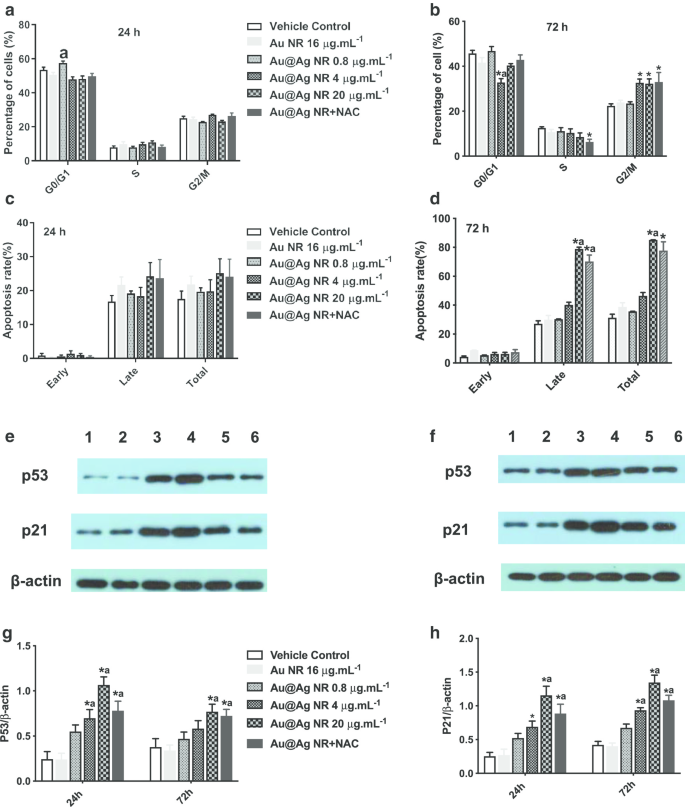
Efectos de Au @ Ag NR sobre el ciclo celular y la apoptosis. Efectos de Au @ Ag NR en el ciclo celular ( a , b ) y apoptosis ( c , d ) después de la exposición durante 24 hy 72 h, respectivamente; los datos representativos de los niveles de expresión de p53 y p21 en células HepaRG de diferentes grupos ( e , f Carril 1:control de vehículos; Carril 2:Au NR; Carril 3:Au @ Ag NR + NAC; Carril 4:Au @ Ag NR 20 μg mL −1 ; Carril 5:Au @ Ag NR 4 μg mL −1 ; Carril 6:Au @ Ag NR 0,8 μg mL −1 ); el nivel de expresión relativa promedio de p53 y p21 a β-actina en diferentes grupos se resumió en ( g , f ). * P <0,05 frente al control del vehículo; a P <0,05 frente a Au NR
Los niveles de expresión de p21 y p53 se detectaron mediante transferencias Western y se observó un patrón similar. Los niveles de expresión de p53 y p21 en células tratadas con 4 μg mL −1 y 20 μg mL −1 Au @ Ag NR aumentaron notablemente ( P <0.05) y disminuyeron significativamente en las células tratadas con 20 μg mL −1 Au @ Ag NR y NAC ( P <0.05, en comparación con 20 μg mL −1 Grupo Au @ Ag NR, Fig. 6e – h). Se sabe que la proteína p53 es una molécula central que media la activación del punto de control G2 / M en respuesta al daño del ADN, y p21 se reconoce como un inhibidor del ciclo celular dependiente de p53. Por lo tanto, el Au @ Ag NR podría interferir con la replicación del ADN y dificultar la reparación del ADN por la detención del ciclo celular.
Discusión
En la actualidad, los roles del Ag + liberado y los AgNP en la generación de genotoxicidad están lejos de ser claros. Estudios previos de nuestro grupo [21] y otros [13] han demostrado que mientras Ag + es la principal fuente de introducción de toxicidades, las nanopartículas también podrían ser altamente tóxicas. Por ejemplo, los AgNP podrían contribuir a la genotoxicidad al inducir la formación de radicales hidroxilo [13]. Además, AgNP introdujo daños cromosómicos más graves, estrés oxidativo y apoptosis en comparación con Ag + solo [23], lo que sugiere que podrían estar involucradas diferentes vías. Empleamos Au @ Ag NR como material modelo para comprender las formas y distribuciones de los AgNP en las células, y las cantidades de Ag y Au intracelulares se determinaron mediante ICP-MS. La relación en peso Ag / Au de Au @ Ag NR preparado se estimó en 2,3. Sin embargo, después de una exposición de 24 h, aumentó bruscamente a 16,5 en las células tratadas con Au @ Ag NR, lo que sugiere que se liberó una gran cantidad de Ag de la cáscara de Au @ Ag NR dentro de ese período. When the exposure period of Au@Ag NR was extended to 72 h, the Au/Ag weight ratio was decreased to 1.7, indicating that the Ag + was released from the cell and the nanorod was the major form of Au@Ag NR in the cell at that stage. Therefore, it could be deduced that once the Au@Ag NR entered the cell, Ag + rapidly dissolved from its shell within 24 h and gradually released to the extracellular environment, while the Au@Ag NR itself retained in the cell for a longer period.
Oxidative stress is deemed as one of the most important toxicological mechanisms of nanoparticles [24]. N-acetylcysteine (NAC) is a thiol, a mucolytic agent and precursor of l-cysteine which reduced glutathione. NAC is also a source of sulfhydryl groups in cells and exerts the ROS scavenger activity by interacting with OH · y H 2 O 2 [25]. In this study, the GSH and SOD levels were significantly decreased after exposure to Au@Ag NR, while the MDA level increased in a concentration- and time-dependent manner, indicating that the Au@Ag NR introduced the oxidative stress in the cells.
The potentials of Ag and Au@Ag NR in interfering with the genetic materials were further investigated by a series of genotoxicity assays. It is noteworthy that co-culturing the NAC with Au@Ag NR could ameliorate the ROS formation, which in turn supports the participation of oxidative stress in the genotoxicity triggered by Au@Ag NR. In this study, comet and γ-H2AX assays were performed to confirm that Au@Ag NR could interact with DNA and induce certain DNA damage, and the repair endonuclease Fpg was included in the comet assay to identify the oxidative DNA damage [26]. The Fgp could recognize oxidized pyrimidines and remove oxidized purines, e.g., 8-hydroguanine, so as to create apurinic or apyrimidinic sites that could introduce gaps in the DNA strands. The oxidative stress-induced DNA breakage could be determined subsequently by another comet assay [27]. The further DNA breakage detected by the additional Fgp in the comet assay suggested that the Au@Ag NR could cause DNA damage. Mei et al. [28] observed that 5-nm-sized AgNPs induced oxidative lesion-specific DNA damage by employing the hOGG1, EndoIII and Fpg endonucleases in the comet assay. Li y col. [29] also suggested that both PVP- and silica-coated AgNPs (15–100 nm and 10–80 nm, respectively) could lead to a significant increase in DNA breakage in mice hepatocytes in the presence of hOGG1and EndoIII. The formation of γ-H2AX foci, which represents an early cellular response to genotoxic stress, is the most sensitive and specific biomarker for detecting DSBs [30]. As demonstrated in this study, γ-H2AX foci in cells exposed to Au@Ag NR were markedly increased after 24 h, and a further increase could be observed after 72 h. The reduction in the 20 µg mL −1 group might be due to the cytotoxicity to the HepaRG cells at higher concentration. Similar results were observed for AgNPs with different coatings [31, 32]. Further, our results suggest that Au@Ag NR could induce chromosome damage in HepaRG cells, as the micronucleus rates were significantly increased. This is consistent with previous studies, where AgNPs-induced increased micronucleus rate was reported in HaCaT and TK6 cells [33]. In contrast, the addition of oxidative radical scavenger NAC could inhibit the formation of micronucleus induced by Au@Ag NR. Taken together, these data suggest the participation of oxidative stress in AgNP-introduced clastogenicity risk in vitro.
Previous studies have investigated the cell cycle arrest and cytotoxicity induced by AgNPs [33,34,35]. With prolonging the exposure time, the impact of AgNPs on cell cycle and apoptosis might be enhanced and in turn aggravate the cytotoxicity and genotoxicity. Usually, the cell cycle checkpoints (e.g., G2/M) were initiated by cells when experiencing DNA damage, and this mechanism serves to prevent the cell from entering mitosis (M phase). The G2/M cell cycle arrest indicates that an increasing percentage of cells is hindered in G2 phase for DNA repairing. Cells experiencing successful DNA repairing would further proceed to mitosis; however, for those with fatal damages, irreversible G2/M cell cycle arrest and cells apoptosis would take place [36]. We observed that Au@Ag NR could arrest the majority of HepaRG cells in G2/M phase, induce late cell apoptosis and increase the expression levels of p53 and p21, which are important proteins associated with the regulation of cell cycles [37]. As p53 could also induce apoptosis, when the DNA cannot be repaired properly [38], the p21 might indirectly participate in cell apoptosis by cell cycle arrest in a p53-dependent pathway via down-regulating the nuclear protein ICBP90 for DNA replication and cell cycle regulation [39]. Furthermore, apoptosis and a G2/M arrest induced by activation of the p53/p21 system have been reported in HepG2 cells following the administration of garlic extracts [40]. Thus, it could be inferred that the oxidative stress-triggered DNA/chromosome damages might facilitate the expression of p53 and p21, which subsequently induces cell cycle arrest. Extending the exposure period of Au@Ag NRs to the DNA/chromosome during replication may further aggravate the genotoxicity or apoptosis.
Conclusion
Genotoxicity induced by AgNPs may be attributed to the oxidative stress induced by the nanoparticles as well as the released ions [41]. This study employed Au@Ag NR as a model to determine the distribution and release behavior of Ag after the nanoparticles enter into the cells. Considering the disparate forms of Au@Ag NR in the cell, after its exposure the Ag + was rapidly dissolved from the silver shell. Ag + and Au@Ag NR could introduce cytotoxicity and genotoxicity (clastogenicity) in the cells, and the Au@Ag NR retained in the nucleus may further release Ag + to aggravate the damage, which are mainly caused by cell cycle arrest and ROS formation (summarized in Fig. 7). Collectively, these data reveal the correlation between the intracellular accumulation, Ag + release as well as the potential genotoxicity of AgNPs.

Schematic diagram of the possible mechanism of genotoxicity introduced by AgNP in vitro
Disponibilidad de datos y materiales
All data and materials are available without restriction.
Nanomateriales
- La diferencia entre la aleación de tungsteno y plata y la aleación de tungsteno y cobre
- Preparación de nanopartículas de mPEG-ICA cargadas con ICA y su aplicación en el tratamiento del daño celular H9c2 inducido por LPS
- Examen de las funciones del tamaño de las gotas de emulsión y del tensioactivo en el proceso de fabricación de nanocristales micelares basado en la inestabilidad interfacial
- Efecto de la distribución de nanopartículas de oro en TiO2 sobre las características ópticas y eléctricas de las células solares sensibilizadas por colorante
- La preparación de la nanoestructura yema-cáscara de Au @ TiO2 y sus aplicaciones para la degradación y detección del azul de metileno
- Promoción del crecimiento celular SH-SY5Y mediante nanopartículas de oro modificadas con 6-mercaptopurina y un péptido penetrante de neuronas
- Profundos efectos interfaciales en CoFe2O4 / Fe3O4 y Fe3O4 / CoFe2O4 Core / Shell Nanoparticles
- Saponinas Platycodon de Platycodi Radix (Platycodon grandiflorum) para la síntesis verde de nanopartículas de oro y plata
- Nanorods de oro modificados con sílice conjugada con anticuerpo para el diagnóstico y la terapia fototérmica de Cryptococcus neoformans:un experimento in vitro
- Evaluación de las propiedades antimicrobianas, apoptóticas y de administración de genes de células cancerosas de nanopartículas de oro cubiertas con proteínas sintetizadas a partir del hongo mic…
- Nanocristales de núcleo / capa de ZnInS / ZnS dopados con Cu sin Cd:Síntesis controlada y propiedades fotofísicas