


El microARN-18b-3p exosómico derivado de células madre mesenquimatosas del cordón umbilical humano inhibe la aparición de preeclampsia al dirigirse a la LEP
Resumen
Los exosomas derivados de células madre mesenquimales del cordón umbilical humano (hucMSC) que expresan microARN se han destacado en enfermedades humanas. Sin embargo, el mecanismo molecular detallado de miR-18b-3p exosómico derivado de hucMSC en la preeclampsia (PE) sigue siendo una investigación adicional. Nuestro objetivo era investigar el efecto de los exosomas y miR-18b-3p / leptina (LEP) sobre la aparición de EP. Se identificó la morfología de los exosomas hucMSC y hucMSC (Exos). Los exosomas se infectaron con diferentes lentivirus que expresan miR-18b-3p para explorar el papel de miR-18b-3p en la PE. El modelo de rata PE se estableció mediante inyección intraperitoneal de N éster metílico de nitro-l-arginina. Se ensayó la expresión de LEP y miR-18b-3p en tejidos de placenta de rata PE. Además, se detectó el efecto de los exosomas sobre la expresión de LEP y miR-18b-3p. Se detectó la presión arterial sistólica (PAS), proteinuria, factores inflamatorios, el peso fetal de rata y placenta y apoptosis celular en ratas PE. Finalmente, la relación entre miR-18b-3p y LEP se verificó mediante el ensayo del gen indicador de luciferasa dual y el ensayo de extracción de ARN. Los exosomas, que restauran miR-18b-3p o inhiben la LEP, redujeron la SBP y la proteinuria de las ratas con PE, así como aumentaron el peso de la rata fetal y la placenta, disminuyeron los niveles séricos de factores inflamatorios y suprimieron las células apoptóticas de las ratas con PE, ejerciendo un efecto supresor. sobre la progresión de la EP. miR-18b-3p disminuyó y la LEP aumentó en los tejidos de la placenta de las ratas PE. LEP fue el gen diana directo de miR-18b-3p. La regulación al alza de miR-18b-3p o el tratamiento de los exosomas suprimieron la expresión de LEP y redujeron la aparición de PE, mientras que la regulación a la baja de miR-18b-3p tuvo efectos contrarios. La LEP regulada negativamente revirtió el efecto de la reducción de miR-18b-3p en ratas PE. El miR-18b-3p exosómico derivado de HucMSC se dirige a la LEP para participar en la aparición y desarrollo de la EP. Este estudio puede proporcionar una base teórica novedosa para el mecanismo y la investigación de la EP.
Introducción
La preeclampsia (EP), caracterizada por proteinuria e hipertensión [1], es una de las principales causas de mortalidad y morbilidad fetal y materna en el embarazo humano [2]. La etiología y patogenia de la EP no están claras [3], que se ha informado que está asociada con una invasión anormal del trofoblasto que resulta en disfunción endotelial materna, mala perfusión placentaria crónica e hipertensión con resultados adversos [4]. Con la excepción del parto fetal y placentario, no existe una terapia específica para la EP [5]. Por tanto, es urgente explorar las dianas terapéuticas para mejorar el pronóstico de esta enfermedad.
El cordón umbilical humano (huc) es una fuente adecuada de células madre mesenquimales (MSC) que secretan una variedad de factores tróficos y citocinas, además de mostrar una fuerte capacidad antiinflamatoria e inmunomoduladora [6]. Un estudio ha verificado que la EP acelera la expresión del marcador neuroglial en las CMM derivadas de la gelatina de Wharton del cordón umbilical [7]. También se ha informado de un efecto protector de los exosomas hucMSCs (Exos) sobre la morfología placentaria y la angiogénesis en ratas PE [8]. Los exosomas son vesículas secretoras pequeñas (50-100 nm) que median la comunicación entre las células en el microambiente del tumor mediante la encapsulación y la transmisión de factores cancerígenos a sitios distantes oa las células circundantes por medio de la circulación [9]. Un estudio ha revelado el daño de las funciones vasculares y las complicaciones inducidas por la transferencia efectiva de sFlt-1 y sEng a las células endoteliales en pacientes con EP por exosomas [10]. Los microARN (miARN) son ARN endógenos, no codificantes, con una longitud de 18-25 nucleótidos, y regulan la expresión de genes a nivel postranscripcional [11]. Los datos de un estudio han informado de que la expresión de miR-18b afecta la invasión celular, la viabilidad y la migración de las células trofoblásticas en la PE [12]. Además, Wu et al. han propuesto que miR-18b atenúa la proliferación en las células endoteliales de la retina humana inducida por niveles altos de glucosa, lo que puede ofrecer una nueva perspectiva para comprender el mecanismo de la patogénesis de la retinopatía diabética [13]. Sin embargo, sigue sin conocerse el papel del miR-18b-3p exosómico derivado de hucMSC en la PE. La leptina (LEP) tiene efectos pleiotrópicos sobre la diferenciación / proliferación celular y la inmunidad de los estados fisiológicos y emergió principalmente de los adipocitos, además de otros tejidos, incluida la placenta [14]. Un estudio ha verificado que la metilación anormal del promotor de la LEP está implicada en la progresión de la EP [15]. Otro estudio ha sugerido que la placenta es un sitio principal de expresión de LEP durante el embarazo [16]. Sin embargo, la relación de unión entre miR-18b-3p y LEP todavía es difícil de alcanzar. Por lo tanto, nuestro objetivo fue explorar el papel de miR-18b-3p exosómico derivado de hucMSC en la EP con la participación de LEP, e inferimos que miR-18b-3p exosómico derivado de hucMSC puede inhibir la progresión de la EP mediante la focalización de la LEP.
Materiales y métodos
Aprobación ética
El estudio fue aprobado por la Junta de Revisión Institucional del Hospital Popular de la Universidad de Wuhan. Todos los participantes firmaron un documento de consentimiento informado. Todos los experimentos con animales coincidieron con la Guía para el cuidado y uso de animales de laboratorio de los Comités Internacionales del Hospital Popular de la Universidad de Wuhan.
Aislamiento, cultivo e identificación de HucMSCs
El cordón umbilical fetal entregado por una puérpera sana se recogió y se cortó en carne picada y se filtró con un tamiz y luego se mezcló con una solución salina tamponada con fosfato (PBS). Los tejidos del cordón umbilical se centrifugaron a 1500 r / min durante 5 min con 10 cm de radio de centrifugación. Los tejidos se suspendieron con medio Eagle modificado de Dulbecco (DMEM) / F12 que contenía suero bovino fetal (FBS) al 10% y se transfirieron a un matraz de cultivo. Los líquidos se cambiaron después de 4 días y luego se cambiaron una vez cada 3 días. Las células se subcultivaron cuando la confluencia alcanzó aproximadamente el 90%. El crecimiento adherente y la morfología de hucMSC se observaron bajo un microscopio óptico. Las células se tiñeron con una solución de tinción de rojo aceite O (Beyotime Institute of Biotechnology, Shanghai, China) para detectar la diferenciación osteogénica de las hucMSC y se tiñeron con una solución de tinción de fosfatasa alcalina (ALP) (Beyotime) para detectar la diferenciación adipogénica de las hucMSC. Se adoptó un citómetro de flujo (Beckman Coulter Life Sciences, Brea, CA, EE. UU.) Para probar CD73, CD166 (ambos 1:10, BD Biosciences, Franklin Lakes, NJ, EE. UU.) Y CD105 (1:20, AbD Serotec, Oxford, Reino Unido).
Extracción e identificación de HucMSC-Exos
Se cultivaron las hucMSC que crecían bien. El sobrenadante se recogió y se centrifugó a 28.500 r / min durante 1 h con un radio de centrifugación de 10 cm. El sobrenadante se descartó y las células se fijaron con glutaraldehído al 2% y ácido ósmico al 1%, se deshidrataron con etanol, se sumergieron en óxido de propileno, se secaron durante 2 h, se incrustaron en Epon812 y se cortaron en rodajas. Las rodajas se tiñeron con uranio y plomo, respectivamente. Finalmente, los exosomas se observaron al microscopio electrónico. Se utilizó un detector de nano visión (Malvern Instruments, Malvern, Reino Unido) para detectar imágenes de movimiento browniano de nanopartículas de exosomas y su tamaño. Los marcadores de superficie de hucMSC-Exos se identificaron mediante un ensayo de transferencia Western y los resultados mostraron que hucMSC-Exos expresaba CD9, CD81 y CD63.
Método de infección por lentivirus
HucMSC se infectó con lentivirus que contenían baja expresión del vector miR-18b-3p y baja expresión del vector control negativo (NC) miR-18b-3p (Shanghai GenePharma Co, Ltd, Shanghai, China). Finalmente, se obtuvieron hucMSC-antagomir NC y hucMSC-miR-18b-3p antagomir expresados de forma estable. Las células se cultivaron durante 48 h, y el sobrenadante se recogió y centrifugó con ultracentrifugación para obtener el correspondiente Exos-antagomir NC y Exos-miR-18b-3p antagomir.
Animales experimentales
Se seleccionaron ratas Wistar (con un peso de 200 a 250 g, un envejecimiento de 8 w, independientemente del sexo) con un nivel de higiene y madurez sexual (el Centro de Animales Experimentales de la Universidad de Wuhan, Wuhan, China). Las ratas fueron alimentadas en un sistema de barrera con una temperatura de 18 a 28 ° C, una humedad relativa de 40 a 70% y una dieta y agua adecuadas.
Establecimiento de modelos de PE de rata
El modelo de PE de rata se estableció mediante inyección intraperitoneal de 50 mg / kg de inhibidor de óxido nítrico sintetasa, éster metílico de N (G) -nitro-1-arginina (L-NAME, Beyotime) con referencia a un artículo [17]. El establecimiento exitoso del modelo de EP se basó en un aumento de la presión arterial con 20 mmgHg y superior a 115 mmHg, así como un aumento de la proteinuria.
Agrupación de animales
La rata hembra y la rata macho cohabitaron aleatoriamente en una proporción de 1:1, y las dos ratas se mantuvieron en una jaula especial individual entre las 5 y las 6 pm. el día anterior. Los espermatozoides de las secreciones vaginales de las ratas hembras se observaron mediante tapón vaginal y microscopio al día siguiente. Si el resultado fue positivo a la misma hora, el día se registró como el día 0 de gestación. Desde el 13 rd día de gestación, las ratas se dividieron en 6 grupos (10 ratas en cada grupo):grupo normal (se inyectó la misma cantidad de solución salina normal por vía intraperitoneal desde el día 13 hasta el día 20 de gestación), grupo PE (L-NAME [50 mg / kg por día] se inyectó intraperitonealmente desde el día 13 hasta el día 20 de gestación, y se inyectaron 20 μL de solución salina normal en la placenta del día 16 al día 19 de gestación), grupo PE + miR-NC (L-NAME [50 mg / kg por día] se inyectó intraperitonealmente desde el día 13 hasta el día 20 de gestación, y se inyectaron 20 μL de 4 nmol de miR-NC en la placenta del día 16 al día 19 de gestación), grupo de PE + miR-18b-3p agomir (Se inyectó L-NAME [50 mg / kg por día] por vía intraperitoneal desde el día 13 hasta el día 20 de gestación, y se inyectaron 20 μL de 4 nmol de miR-18b-3p agomir en la placenta del día 16 al día 19 de gestación) , El grupo de PE + miR-18b-3p antagomir (L-NAME [50 mg / kg por día] se inyectó por vía intraperitoneal desde el día 13 hasta el día 20 de gestación, y se inyectaron 20 μL de 4 nmol de miR-18b-3p antagomir en la pags lacenta en el día 16 al día 19 de gestación), PE + miR-18b-3p antagomir + grupo de ARN interferente pequeño (si) -LEP (L-NAME [50 mg / kg por día] se inyectó por vía intraperitoneal desde el día 13 al día 20 de gestación, y se inyectaron 20 μL de 4 nmol de miR-18b-3p antagomir y si-LEP en la placenta del día 16 al día 19 de gestación) y al grupo de PE + si-LEP (L-NAME [50 mg / kg por día] se inyectó intraperitonealmente desde el día 13 al día 20 de gestación, y se inyectaron 20 μL de 4 nmol de si-LEP en la placenta del día 16 al día 19 de gestación). Las ratas se trataron con exosomas y exosomas que portaban lentivirus. Las ratas se asignaron en 5 grupos (10 ratas en cada grupo):grupo normal (se inyectó la misma cantidad de solución salina normal por vía intraperitoneal desde el día 13 hasta el día 20 de gestación), grupo PE (L-NAME (50 mg / kg por día) ) se inyectó por vía intraperitoneal desde el día 13 hasta el día 20 de gestación, y se inyectaron 20 μL de solución salina normal en la placenta del día 16 al día 19 de gestación), grupo PE + Exos (L-NAME (50 mg / kg por día) se inyectó por vía intraperitoneal desde el día 13 hasta el día 20 de gestación, y se inyectaron 20 μL de Exos (se suspendieron 80 μg de exosomas en 20 μL de solución salina normal) en la placenta del día 16 al día 19 de gestación), PE + Exos-antagomir NC grupo (L-NAME (50 mg / kg por día) se inyectó intraperitonealmente desde el día 13 hasta el día 20 de gestación, y se inyectaron 20 μL de Exos-antagomir NC (80 μg de exosomas se suspendieron en 20 μL de solución salina normal) en la placenta del día 16 al día 19 de gestación) y el grupo de PE + Exos-miR-18b-3p antagomir (L-NAME (50 mg / kg por día) se inyectó por vía intraperitoneal desde el día 13 hasta el día 20 de gestación, y se inyectaron 20 μL de Exos-miR-18b-3p antagomir (se suspendieron 80 μg de exosomas en 20 μL de solución salina normal) en la placenta entre el día 16 y el día 19 de gestación).
Detección de presión arterial sistólica (PAS) y determinación de proteinuria de 24 h
La presión de las ratas se midió mediante la medición de la presión sanguínea de la arteria de la cola de las ratas. La PAS del manguito de la cola de todas las ratas preñadas se midió en los días 10, 13, 16 y 19 de gestación utilizando el detector de presión de la arteria de la cola de rata (Tensys (R) Medical Inc., San Diego, CA, EE. UU.). La presión se midió 3 veces en poco tiempo; luego, el valor promedio se tomó como la presión arterial.
En el caso de la dieta y el agua libres, la orina de 24 h de ratas preñadas se recogió los días 10, 13, 16 y 19 de la gestación, y el contenido de proteínas se detectó en el departamento de nefrología del Hospital Popular de la Universidad de Wuhan.
Colección de muestra
Las ratas preñadas se anestesiaron con pentobarbital sódico al 3% el día 21 de gestación. La sangre periférica de las ratas se conservó, se centrifugó para tomar el suero y se almacenó en el frigorífico a -20 ° C como espera. Luego, la rata fetal y la placenta se extrajeron mediante cesárea, se extrajeron la membrana fetal y el cordón umbilical conectado, y se cortó el cordón umbilical conectado a la rata fetal. La placenta y la rata fetal se colocaron sobre una gasa aséptica para secar la sangre y el líquido amniótico, respectivamente, y luego se colocaron en la balanza analítica para pesar el peso. Una parte de los tejidos placentarios se fijó con paraformaldehído al 4%, se deshidrató con etanol, se aclaró con xileno, se incrustó con parafina y se cortó continuamente (5 μm) para tinción con hematoxilina-eosina (HE) y desoxiuridina trifosfato-biotina mediada por desoxiuridina trifosfato terminal. tinción de etiquetado de extremos de muesca (TUNEL). El resto se almacenó a -80 ° C para la detección de la reacción en cadena de la polimerasa cuantitativa con transcripción inversa (RT-qPCR), el análisis de transferencia Western y el ensayo inmunoabsorbente ligado a enzimas (ELISA).
ELISA
Los contenidos de factor de necrosis tumoral-α (TNF-α), interleucina (IL) -1β e IL-6 en suero se analizaron mediante ELISA. Las concentraciones de TNF-α, IL-1β e IL-6 se determinaron siguiendo las instrucciones del kit (R&D Systems, Minneapolis, MN, EE. UU.). Los valores de densidad óptica (DO) (490 nm) se probaron con un lector de microplacas (Thermo Fisher Scientific, MA, EE. UU.). La curva estándar correspondiente se obtuvo usando el valor de DO como abscisa y la concentración de la muestra estándar correspondiente como ordenada. Las concentraciones de TNF-α, IL-1β e IL-6 se calcularon a partir de la curva estándar.
Tinción HE
Las muestras de parafina de los tejidos de la placenta se aclararon en xileno, se deshidrataron con alcohol en gradiente convencional, se tiñeron con hematoxilina, se diferenciaron con alcohol ácido clorhídrico al 1% y se volvieron a azul con agua amoniacal al 1%. Luego, los tejidos se tiñeron con una solución de eosina al 1%, se deshidrataron (75%, 90%, 95% de etanol, respectivamente, alcohol etílico absoluto) y se aclararon con xileno, se secaron, se bloquearon y se observaron al microscopio electrónico.
Tinción TUNEL
Las secciones incluidas en parafina se desparafinaron y deshidrataron rutinariamente de acuerdo con las instrucciones, y luego, se detectó la apoptosis con el kit TUNEL (Nanjing Kejin Biotechnology Co., Ltd., Jiangsu, China). Se usó 4,6-diamino-2-fenilindol (Shanghai Baitai Biotechnology Co., Ltd., Shanghai, China) para observar células TUNEL positivas usando un microscopio de fluorescencia (Nikon, Tokio, Japón) [18].
RT-qPCR
Se pesaron los tejidos de la placenta. Por cada 50-100 mg de tejido de placenta se añadió 1 ml de TRIzol (Invitrogen, Carlsbad, California, EE. UU.) Y se disolvió por completo. Los tejidos se agregaron con 200 μL de cloroformo y se centrifugaron a 4 ° C, 12.000 rpm para extraer el ARN total. La concentración y pureza del ARN se determinaron mediante espectrofotómetro de ácidos nucleicos de proteínas DU-800 (Beckman). Se utilizaron U6 y β-actina como controles de carga. Los cebadores de PCR fueron diseñados y compuestos por Shanghai Sangon Biotechnology Co. Ltd. (Shanghai, China). Las secuencias de los cebadores se enumeran en la Tabla 1. El ARN se invirtió en ADNc sobre la base de las instrucciones del kit de transcripción inversa de ARN (Sangon). Se amplificó la PCR y los productos se verificaron mediante electroforesis en gel de agarosa. Los datos fueron calculados por 2 −ΔΔCt método.
Ensayo de transferencia Western
La proteína total de los tejidos de la placenta se extrajo mediante tampón de lisis celular de ensayo de radioinmunoprecipitación (Beyotime). Se utilizó HucMSC-Exo para abstraer el tampón, que se centrifugó a 14.000 rpm. El sobrenadante se conservó para probar la expresión proteica de la proteína marcadora exosómica (CD81, CD63 y CD9) en suero. La concentración de proteína se determinó mediante el kit de ácido bicinconínico (Beyotime, P0010). La muestra se cargó de acuerdo con los resultados cuantitativos de proteína, se trató con electroforesis en gel de poliacrilamida-dodecilsulfato de sodio al 10% y se transfirió a la membrana. La membrana se bloqueó con leche desnatada al 5%, se sondaron con anticuerpos primarios LEP, CD63, CD81, CD9 y β-actina (4 mL, 1:1000, Santa Cruz Biotechnology, Inc, Santa Cruz, CA, EE. UU.), Se volvieron a sondar con 4 mL de anticuerpo secundario de cabra anti-conejo IgG / peroxidasa de rábano picante, expuestos y revelados. Se utilizó β-actina como referencia interna. El valor de gris se analizó mediante el software de análisis gráfico de gel Image Lab.
Ensayo génico indicador de luciferasa dual
Se adoptó el software de predicción en línea https://cm.jefferson.edu/ para predecir la relación objetivo entre miR-18b-3p y LEP, así como el sitio de unión de miR-18b-3p y la región no traducida de LEP 3 '(UTR ). Se compuso la secuencia de la región promotora 3'UTR de LEP que contenía el sitio de unión de miR-18b-3p. Se construyeron el plásmido de tipo salvaje LEP 3'UTR (WT) y el tipo mutante LEP 3'UTR (MUT). Los plásmidos recombinantes se denominaron LEP 3'UTR-WT y LEP 3'UTR-MUT, respectivamente. Las células 293T cultivadas se cotransfectaron con míR-18b-3p mimic y LEP 3'UTR-WT, miR-18b-3p mimic y LEP 3'UTR-MUT, imitan NC y LEP 3'UTR-WT, imitan NC y LEP 3′UTR-MUT durante 30 h. Luego se recolectaron células 293T. La actividad luciferasa de luciérnaga y renilla en las células se midió mediante mediciones de luminiscencia de acuerdo con el kit de detección del gen indicador de luciferasa dual (Promega, Madison, WI, EE. UU.).
Ensayo de extracción de ARN
Se incubaron sondas de ARN biotinilado (Bio-miR-NC, Bio-miR-18b-3p y Bio-miR-18b-3p-Mut) con el lisado de células 293T y se extrajeron usando perlas magnéticas conjugadas con antibiótico estreptomicina. El experimento se realizó según las instrucciones de los kits de extracción de ARN magnético de Pierce (Pierce, IL, EE. UU.). El ARN se eluyó y purificó usando TRIzol (Pierce). El enriquecimiento de LEP en el complejo de ARN se cuantificó utilizando RT-qPCR como se describió anteriormente [19].
Análisis estadístico
Todos los datos fueron explicados por el software SPSS 21.0 (IBM Corp. Armonk, NY, EE. UU.). Los datos de medición se indicaron como media ± desviación estándar. Los datos fueron obtenidos por una muestra independiente t prueba para las comparaciones de dos grupos, mientras que las comparaciones entre múltiples grupos se evaluaron mediante un análisis de varianza de una vía (ANOVA) seguido de la prueba post hoc de Tukey. El criterio de significación estadística se estableció en p <0.05.
Resultados
Morfología e identificación de HucMSC y HucMSC-Exos
Las masas de tejido del cordón umbilical se observaron bajo un microscopio invertido. Se pudo ver que las células salieron de la masa de tejido el día 3; las células mostraban forma de huso y filiformes y crecían como una colonia aproximadamente a los 5 días. Cuando se cultivaron hasta el paso 3, la morfología de las células fue uniforme, fusiforme larga y similar a la morfología de los fibroblastos, y la disposición fue regular (Fig. 1a). Después de 2 w de diferenciación adipogénica de hucMSC, se formaron gotitas de lípidos en el citoplasma, y las gotitas de lípidos mostraron estructura de Kranz bajo el microscopio invertido (Fig. 1b), lo que sugiere que las hucMSC cultivadas aisladas tenían la capacidad de diferenciación adipogénica. Después de 2 w de diferenciación osteogénica, se pudo ver un gran número de nódulos de calcio marrón bajo un microscopio invertido (Fig. 1c), lo que indica que las hucMSC cultivadas aisladas tenían la capacidad de diferenciación osteogénica. Se adoptó un citómetro de flujo para probar el inmunofenotipo de las células, y los resultados incluyeron que las células sobreexpresaban los marcadores de superficie CD73, CD105 y CD166 de las MSC (Fig. 1d).
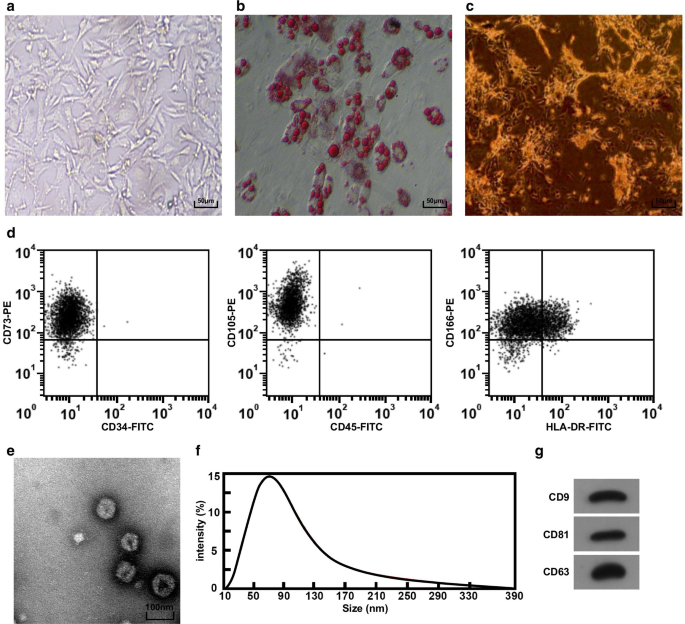
Morfología e identificación de hucMSC y hucMSC-Exos. un La morfología del hucMSC bajo el microscopio invertido, b hucMSC se ensayó mediante tinción con rojo aceite O. c hucMSC se ensayó mediante tinción ALP. d Se utilizó citometría de flujo para detectar inmunofenotipo. e La forma y el tamaño de hucMSC-Exos observados a través de un TEM. f Detección de la distribución del tamaño de partícula de exosomas mediante análisis Nanosight. g La expresión de proteínas de CD9, CD81 y CD63 en hucMSC-Exos se detectó mediante un ensayo de transferencia Western
La morfología de hucMSC-Exos fue observada por el TEM, y los resultados presentaron que los exosomas eran redondos u ovalados con baja densidad central y tinción espesa en ambos lados (Fig. 1e). Se utilizó análisis de nano-visión para analizar el tamaño de partícula de exosomas, y los resultados mostraron que el tamaño de partícula se distribuyó principalmente entre 40 y 100 nm, más concentrado alrededor de 80 nm (Fig. 1f). El ensayo de transferencia Western reveló que todos los marcadores de superficie CD81, CD63 y CD9 se expresaron en hucMSC-Exos (Fig. 1g).
La restauración de miR-18b-3p alivia las características patológicas de las ratas PE
Los resultados de PAS y proteinuria de 24 h presentaron que:no hubo diferencia significativa en PAS y proteinuria de 24 h en 6 grupos antes de la administración (día 10 de gestación). La PAS y la proteinuria de 24 h en el día 19 de gestación no mostraron diferencias obvias en ratas normales. En ratas PE o ratas PE tratadas con miR-NC, miR-18b-3p agomir, miR-18b-3p antagomir, miR-18b-3p antagomir + si-LEP, si-NC o si-LEP, SBP y 24-h La proteinuria comenzó a aumentar a los 13 días de gestación. No hubo diferencia clara de SBP y proteinuria de 24 h en el día 16 y el día 19 de gestación en ratas PE tratadas con miR-18b-3p agomir y si-LEP. Las ratas PE habían aumentado la PAS y la proteinuria de 24 h en el día 19 de gestación; este aumento se redujo por la elevación de miR-18b-3p, pero mejoró aún más por la inhibición de miR-18b-3p; La reducción de LEP anuló el papel de la regulación a la baja de miR-18b-3p en la PAS y la proteinuria de 24 h en el día 19 de gestación en ratas PE (Fig. 2a, b).
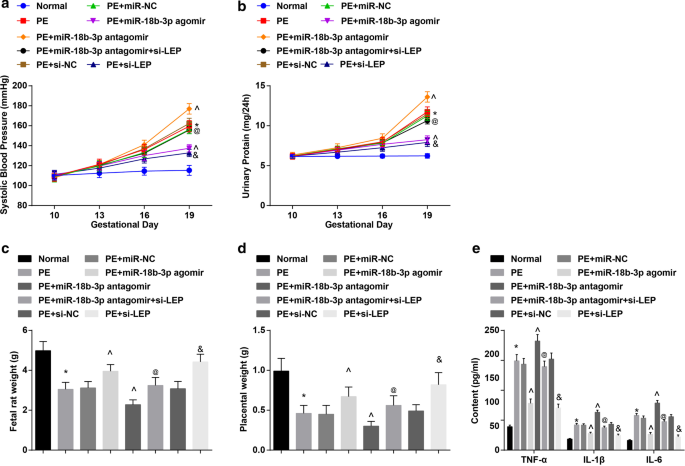
La restauración de miR-18b-3p alivia las características patológicas de las ratas PE. un Resultados de la PAS en ratas. b Resultados de la proteinuria de 24 h en ratas. c Cambios de peso de ratas fetales. d Cambios de peso placentario en ratas. e Los cambios de factores inflamatorios en suero se detectaron mediante ELISA. n =10, * p <0,05 frente al grupo normal. ^ p <0,05 frente al grupo PE + miR-NC. @ p <0,05 frente al grupo de antagomir PE + miR-18b-3p. y p <0,05 frente al grupo PE + si-NC. Los datos de medición se representaron como media ± desviación estándar, y las comparaciones entre varios grupos se evaluaron mediante ANOVA de una vía seguida de la prueba de Tukey
El peso de la rata fetal y la placenta se redujeron en las ratas PE; miR-18b-3p regulado al alza o LEP regulado a la baja aumentó, mientras que miR-18b-3p regulado a la baja disminuyó el peso de la rata fetal y la placenta en ratas PE; El silenciamiento de LEP revirtió el efecto de la inhibición de miR-18b-3p sobre el peso de la rata fetal y la placenta en ratas PE (Fig. 2c, d).
Se detectaron factores inflamatorios en suero de ratas PE. Se encontró que los contenidos de TNF-α, IL-1β e IL-6 aumentaron en las ratas PE; Se suprimió la elevación de miR-18b-3p o la inhibición de LEP, mientras que la reducción de miR-18b-3p promovió los contenidos de TNF-α, IL-1β e IL-6; el efecto de miR-18b-3p inhibido sobre los contenidos de TNF-α, IL-1β e IL-6 fue anulado por la disminución de LEP (Fig. 2e).
El miR-18b-3p sobreexpresado mejora el cambio histopatológico de los tejidos placentarios de ratas con EP
En ratas normales, las vellosidades placentarias eran ricas en vasos sanguíneos y tenían una estructura clara, los sincitiotrofoblastos eran los principales trofoblastos en las vellosidades placentarias y había menos citotrofoblastos. En ratas PE o ratas PE tratadas con miR-NC, miR-18b-3p antagomir, si-NC o miR-18b-3p antagomir + si-LEP, el número de vellosidades placentarias disminuyó, la estructura se volvió borrosa y atrofiada, algunas vellosidades se realizaron necrosis fibrinoide, y aumentó el número de nódulos sincitiotrofoblasto en las vellosidades placentarias, y la mayoría de las vellosidades eran inmaduras. Se redujo el número de trofocitos y se aliviaron los cambios patológicos en ratas PE tratadas con miR-18b-3p agomir y si-LEP (Fig. 3a).
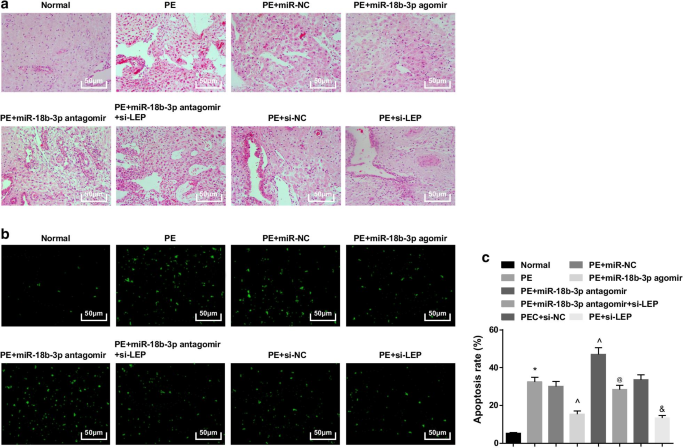
El miR-18b-3p sobreexpresado mejora el cambio patológico y suprime las células apoptóticas de los tejidos de la placenta en ratas PE. un La tinción HE se utilizó para probar las características patológicas de los tejidos de la placenta. b La tinción TUNEL se implementó para determinar las células apoptóticas de los tejidos de la placenta en ratas PE. c La tasa de apoptosis celular se detectó mediante tinción TUNEL. n =10, * p <0,05 frente al grupo normal. ^ p <0,05 frente al grupo miR-NC. @ p <0,05 frente al grupo de miR-18b-3p antagomir. y p <0,05 frente al grupo si-NC. Los datos de medición se representaron como media ± desviación estándar, y las comparaciones entre varios grupos se evaluaron mediante ANOVA de una vía seguida de la prueba de Tukey
La tinción TUNEL sugirió que podía verse una pequeña cantidad de células apoptóticas. Las ratas PE habían aumentado las células apoptóticas, que se redujeron por la elevación de miR-18b-3p y el silenciamiento de LEP, y fueron mejoradas aún más por la inhibición de miR-18b-3p; El silenciamiento de LEP también revirtió el efecto de la inhibición de miR-18b-3p sobre el número de células apoptóticas en ratas PE (Fig. 3b, c).
En conjunto, las ratas con miR-18b-3p regulado al alza o LEP inhibida tenían un grado reducido de progresión de la EP en histología, y la LEP silenciada podría abolir el efecto terapéutico del miR-18b-3p inhibido.
miR-18b-3p está regulado a la baja, mientras que LEP está regulado al alza en los tejidos de placenta de rata con PE y miR-18b-3p se dirige a LEP
En base a los resultados anteriores, la regulación a la baja de LEP revirtió el efecto terapéutico de la regulación a la baja de miR-18b-3p en ratas PE en patología e histología; por lo tanto, planteamos la hipótesis de que miR-18b-3p puede estar relacionado con LEP.
El ensayo de transferencia Western y RT-qPCR revelaron que las ratas PE habían disminuido miR-18b-3p y habían aumentado los niveles de expresión de LEP; el tratamiento de miR-18b-3p agomir regulaba positivamente miR-18b-3p y regulaba negativamente la LEP en ratas PE, mientras que el tratamiento de miR-18b-3p antagomir aumentaba la expresión de LEP; El silenciamiento de LEP revirtió el efecto promotor de la reducción de miR-18b-3p sobre la expresión de LEP en ratas PE (Fig. 4a-c).
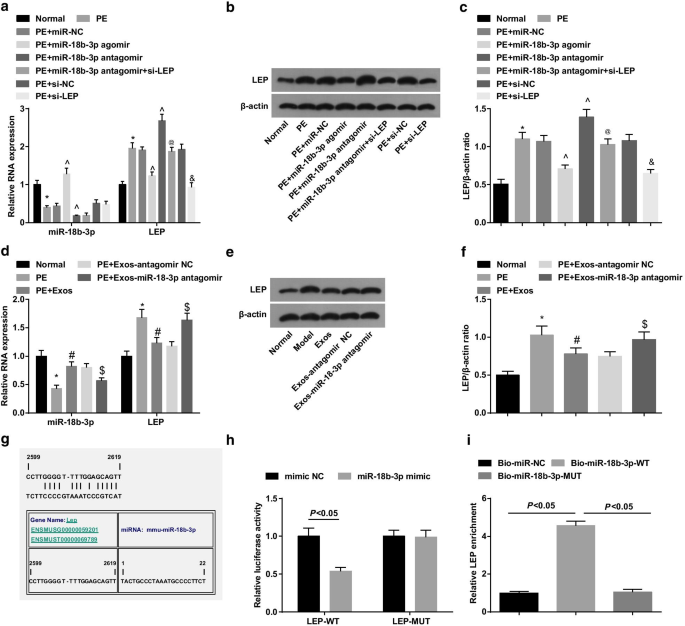
miR-18b-3p se regula a la baja y la LEP se regula al alza en los tejidos de la placenta de las ratas PE. un La expresión de ARNm de miR-18b-3p y LEP en tejidos de placenta se detectó usando RT-qPCR. b Banda proteica de LEP en tejidos placentarios. c Se llevó a cabo un ensayo de transferencia Western para detectar la expresión de la proteína LEP en los tejidos de la placenta. d Se detectó la expresión de ARNm de miR-18b-3p y LEP en tejidos de placenta después del tratamiento con exosomas usando RT-qPCR. e Banda de proteína de LEP en tejidos de placenta después del tratamiento con exosomas. f Se realizó un ensayo de transferencia Western para detectar la expresión de la proteína LEP después del tratamiento con exosomas. g Los sitios de unión de miR-18b-3p y LEP predicados por un software en línea. h La relación diana entre miR-18b-3p y LEP verificada mediante el ensayo del gen indicador de luciferasa dual. yo Relación de focalización entre miR-18b-3p y LEP verificada por ensayo de reducción de ARN. n =10, * p <0,05 frente al grupo normal. ^ p <0,05 frente al grupo miR-NC. @ p <0,05 frente al grupo de miR-18b-3p antagomir. y p <0,05 frente al grupo si-NC. # p <0,05 frente al grupo de EP. $ p <0,05 frente al grupo PE + Exos-antagomir NC. Los datos de medición se representaron como media ± desviación estándar, y las comparaciones entre varios grupos se evaluaron mediante ANOVA de una vía seguida de la prueba de Tukey
Se utilizaron el ensayo de transferencia Western y RT-qPCR para explorar el papel de los exosomas en ratas PE. Los resultados mostraron que los exosomas aumentaron miR-18b-3p y disminuyeron la LEP en ratas PE, lo que indica el efecto supresor de los exosomas en el desarrollo de PE. Además, los exosomas que transportan miR-18b-3p antagomir indujeron la regulación a la baja de miR-18b-3p y la regulación al alza de LEP en ratas PE (Fig. 4d-f).
La relación objetivo entre miR-18b-3p y LEP fue pronosticada por el software de predicción bioinformática en línea https://cm.jefferson.edu/ (Fig. 4g). El ensayo del gen indicador de luciferasa dual sugirió que el mimetismo de miR-18b-3p disminuyó la actividad de luciferasa de LEP 3'UTR-WT, mientras que no impuso impactos sobre la de LEP 3'UTR-MUT (Fig. 4h). Además, el ensayo de reducción de ARN reveló que el enriquecimiento de LEP aumentó mediante miR-18b-3p biotinilado con WT (Fig. 4i). Estos hallazgos indicaron que LEP es un gen diana de miR-18b-3p.
hucMSC-Exos atenúan las características patológicas de las ratas PE
Los resultados de PAS y 24 h presentaron que no hubo diferencia significativa en PAS y proteinuria de 24 h en 5 grupos antes de la administración (día 10 de gestación). SBP and 24-h proteinuria in day 19 of gestation showed no distinct difference in normal rats. In PE rats, SBP and 24-h proteinuria began to raise at day 13 of gestation. There was no distinct difference of SBP and 24-h proteinuria in day 16 and day 19 of gestation in PE rats treated with hucMSC-Exos and hucMSC-Exos transmitting antagomir NC. SBP and 24-h proteinuria heightened in day 19 of gestation in the PE rats, while the increase was reduced by injection of hucMSC-Exos. Inhibiting miR-18b-3p reversed the effect of hucMSC-Exos on SBP and 24-h proteinuria in day 19 of gestation in PE rats (Fig. 5a, b).

hucMSC-Exos attenuate pathological characteristics of PE rats. un Results of SBP in rats after exosome treatment. b Results of 24-h proteinuria in rats after exosome treatment. c Weight changes of fetal rats after exosome treatment. d Changes of placental weight in rats after exosome treatment. e Changes of inflammation factors after exosome treatment in serum were determined using ELISA. n = 10, *p < 0.05 versus the normal group. # p < 0.05 versus the PE group. $ p < 0.05 versus the PE + Exos-antagomir NC group. Measurement data were depicted as mean ± standard deviation, and comparisons among multiple groups were assessed by one-way ANOVA followed by Tukey’s test
The weight of fetal rat and placenta was measured, and we found that the PE rats had decreased weight of fetal rat and placenta; miR-18b-3p downregulation abolished the role of hucMSC-Exos in the weight of fetal rat and placenta in PE rats (Fig. 5c, d).
Inflammatory factors in serum were detected using ELISA. TNF-α, IL-1β and IL-6 contents remarkably increased in PE rats. Exosomes treatment decreased TNF-α, IL-1β and IL-6 contents in serum of PE rats, which were enhanced by injection of exosomes inhibiting miR-18b-3p (Fig. 5e).
Exosomes Alleviates Pathological Change and Inhibit Apoptosis of Placenta Tissues of PE Rats
In normal rats, the placental villus was abundant in blood vessels with a clear structure, syncytiotrophoblasts were the main trophoblast in placental villi, and there were fewer cytotrophoblasts. In the PE rats and PE rats treated with hucMSC-Exos-miR-18b-3p-antagomir, the number of placental villi reduced, the structure was blurred and atrophied, some villi were presented fibrinoid necrosis, and the number of syncytiotrophoblast nodules in placental villi enhanced, and most of the villi were immature. The pathological change was improved in the PE rats treated with hucMSC-Exos or hucMSC-Exos-antagomir NC versus the PE rats and PE rats treated with hucMSC-Exos-miR-18b-3p antagomir (Fig. 6a).

Exosomes alleviate pathological change and decrease apoptotic cells of placenta tissues in PE rats. un HE staining was utilized to test pathological features of placenta tissues in PE rats after exosome treatment. b TUNEL staining was implemented to determine apoptotic cells of placenta tissues in PE rats after exosome treatment. c Cell apoptosis rate was detected by TUNEL staining. n = 10, *p < 0.05 versus the normal group. # p < 0.05 versus the PE group. $ p < 0.05 versus the PE + Exos-antagomir NC group. Measurement data were depicted as mean ± standard deviation, and comparisons among multiple groups were assessed by one-way ANOVA followed by Tukey’s test
TUNEL staining indicated that in normal rats, a small number of apoptotic cells could be seen. PE rats had enhanced apoptotic cells, and reduced miR-18b-3p reversed the impacts of hucMSC-Exos on the number of apoptotic cells in placenta tissues from PE rats (Fig. 6b, c).
Discussion
PE is a multisystem pregnancy disorder characterized by proteinuria and either high blood pressure or other adverse conditions and is linked to a wide range of maternal endothelial dysfunction [20]. It was reported that hucMSC-Exo improved the morphology of placental tissue in PE rats through suppressing cell apoptosis and facilitating angiogenesis in placental tissue in a dose-dependent manner [8]. A study has reported that miR-18b expression affected cell migration, viability and invasion in PE [12]. Moreover, it was verified increased maternal LEP concentration and hypomethylation of the LEP in placenta in early onset PE [21]. The current study was designed to explore the effect of exosomes and miR-18b-3p targeted LEP on the occurrence of PE. The findings in this study revealed that hucMSC-derived exosomal miR-18b-3p inhibited PE progression by reducing LEP.
Based on our findings, miR-18b-3p reduced and LEP elevated in placenta tissues of PE rats. Similar to our study, the mRNA expression of miR-18b was markedly suppressed in PE placental tissues relative to that in normal placental tissues [12]. In addition, a study revealed that miR-18b content was dramatically reduced in malignant melanoma tissues in comparison with their matched adjacent non-tumor tissues [22]. Another study has verified that placental LEP expression was raised in preterm PE compared with controls [23]. Moreover, a study showed that LEP expression was obviously heightened in preeclamptic placentas [15]. This literature provided a theoretical basis for us to explore the abnormal expression of miR-18b-3p and LEP in PE. Moreover, it was predicted using a bioinformatic software that LEP was targeted by miR-18b-3p, and this targeting relationship was further confirmed with dual-luciferase reporter gene assay in our research. A study reported that LEP is a target for all three miRNAs (miR-1301, miR-223 and miR-224) in early-onset PE [16]. Another study has displayed that LEP decreased miR-93 expression in osteoarthritis and rheumatoid arthritis [24]. However, the binding between miR-18b-3p and LEP in human diseases, especially in PE, remains scarcely studied, which is the novelty of this study. Furthermore, a result emerging from our study reported that exosomes increased miR-18b-3p and decreased LEP in placenta tissues of PE. It was formerly documented that the expression of miR-18b-5p was notably raised in colorectal cancer plasma exosomes [25], while the relationship between hucMSC-Exos and miR-18b-3p/LEP in PE needs further study.
Additionally, the finding from our investigation showed that restored miR-18b-3p reduced SBP and 24-h proteinuria of PE rats, increased the weight of placenta, declined TNF-α, IL-1β and IL-6 contents in serum and placenta tissues as well as suppressed cell apoptosis. These data indicated that miR-18b-3p elevation contributes to alleviating the symptoms and pathological changes in PE. It was demonstrated that stable upregulation of miR-18b produced effective tumor inhibitor activity, such as inhibiting melanoma cell viability, inducing apoptosis and reducing tumor growth in vivo [26]. Another result in our study was that hucMSC-Exos reduced SBP and 24-h proteinuria of PE rats, increased the weight of placenta, declined TNF-α, IL-1β and IL-6 contents in serum and placenta tissues as well as suppressed cell apoptosis. The findings of the current study revealed that exosomes treated PE rat models presented an increase of the number and quality of fetuses, the quality of placenta, but cell apoptosis was significantly reduced [8]. Interestingly, a previous research has demonstrated that the addition of fetal bovine exosomes declined contents of macrophage TNF-α and IL-6 [27]. A study has revealed that purified exosomes suppressed production of IL-1β in lipopolysaccharide/nigericin-stimulated macrophages [28]. Furthermore, Nong et al. have suggested that inflammatory markers, such as TNF-α and IL-6, were dramatically decreased after administration of exosomes produced through human-induced pluripotent stem cell-derived MSCs [29]. There is a article finding that the SBP was markedly elevated in the group of women who later developed PE [30, 31]. It was displayed that PE patients were positively associated with SBP and diastolic blood pressures and proteinuria [32]. Also, a recent study has provided a proof that proteinuria heightened with advancing gestation in PE women [33]. A important finding was that rats from the PE group had increased TNF-α relative to the normal pregnant group [34]. Another study has verified that serum IL-6 and IL-1β were obviously elevated in women with PE in relation to controls [35]. The above findings suggested that PE patients usually showed high SBP, proteinuria and levels of inflammatory factors. Thus, it could be inferred from our results that the hucMSC-derived exosomal miR-18b-3p had a therapeutic effect on PE.
Conclusion
In conclusion, our study provides evidence that hucMSCs-derived exosomes upregulate miR-18b-3p, which targets LEP to suppress the contents of inflammatory factors and reduce cell apoptosis rate in PE rat placenta tissues, thereby inhibiting the occurrence of PE. Thus, exosomal miR-18b-3p may be a potential candidate for treatment of PE via targeting LEP. This research identified the role of hucMSC-derived exosomal miR-18b-3p targeting LEP during PE development for the first time, which provided a novel insight for PE treatment. However, due to the limitation of known researches, the study needs to be monitored rigorously and reported appropriately in the future clinical trials.
Disponibilidad de datos y materiales
No aplica.
Nanomateriales
- ¿Cómo daña el cadmio al cuerpo humano?
- Por qué el elemento humano de confiabilidad es tan importante
- Las neuronas artificiales podrían ser tan eficientes como el cerebro humano
- La llegada de la cadena de suministro "autónoma"
- El elemento humano crítico en la estrategia de ciberseguridad
- En la pandemia, el compromiso de las fábricas con los derechos humanos está disminuyendo
- El futuro de la automatización en la industria de la aviación
- Sistema de seguridad biométrico basado en el latido del corazón humano
- Delaware:la combinación perfecta entre humanos e inteligencia artificial
- El elemento humano de la automatización
- El factor humano en la inspección de metales