


Evaluación de la actividad del citocromo P450 3A4 inhibido por nanopartículas de oro y los mecanismos moleculares que subyacen a su toxicidad celular en la línea celular de carcinoma hepatocelular humano C3A
Resumen
Las interacciones de las nanopartículas de oro de 40 y 80 nm (AuNP) funcionalizadas con polietilenimina ramificada catiónica (BPEI), ácido lipoico aniónico (LA) o polietilenglicol neutro (PEG) con la línea celular de carcinoma hepatocelular humano (HCC) C3A se han investigado en el ausencia y presencia de corona de proteínas plasmáticas humanas (PC). Todas las AuNP desnudas (sin PC) además de LA-AuNP de 80 nm fueron citotóxicas para C3A pero la PC atenuó sus citotoxicidades. La captación celular dependiente del tiempo de AuNP aumentó además de BPEI-AuNP de 40 nm, pero la PC suprimió sus captaciones además de PEG-AuNP de 80 nm. Las respuestas bifásicas de estrés oxidativo / nitrosativo por BPEI-AuNP ocurrieron en células C3A, mientras que PEG-AuNP fue un potente antioxidante. Todas las AuNP desnudas inhibieron la actividad del citocromo P450 (CYP) 3A4 independientemente del tamaño y la carga superficial, pero la PC recuperó su actividad además de la PEG-AuNP. La expresión génica modulada por PEG-AuNP de 40 nm participó principalmente en la β-oxidación de ácidos grasos mitocondriales y, en menor grado, en los transportadores de eflujo / captación hepáticos. Estos estudios contribuyen a una mejor comprensión de la interacción AuNP con procesos biológicos clave y sus mecanismos moleculares subyacentes en el HCC, que pueden estar más implicados en el desarrollo de una diana terapéutica más eficaz en el tratamiento del HCC.
Antecedentes
El carcinoma hepatocelular (CHC) es uno de los cánceres más comunes en todo el mundo y la causa de mortalidad por cáncer de más rápido crecimiento en los EE. UU. [1, 2]. Dado que el HCC se ha diagnosticado en estadios avanzados, los tratamientos curativos del HCC incluyen trasplante de hígado o resección quirúrgica en el desarrollo temprano del tumor y quimioterapia y radioterapia para un estado avanzado del tumor. El HCC a menudo desarrolla una alta resistencia a los agentes antineoplásicos convencionales, una molécula citotóxica no selectiva que podría producir efectos adversos sistémicos. Los avances recientes en la terapia génica, es decir, la terapia génica basada en la interferencia de ARN (ARNi), se han utilizado en el tratamiento actual del HCC [3, 4]. La eficacia de RNAi requiere que el vector se administre al interior de la célula diana [5]. Los vectores para una entrega exitosa de genes son vectores virales y no virales. Los virus ofrecen una mayor eficiencia en la entrega de genes, pero se prefieren los vectores no virales debido a problemas de seguridad con los vectores virales. Las nanopartículas (NP) como vectores no virales para la administración de genes dirigida o el sistema de administración de fármacos han ganado una gran atención para mejorar la eficiencia terapéutica y reducir la toxicidad en los niveles sistémicos y / o celulares en el tratamiento del HCC [4, 6]. Por lo tanto, se vuelve muy importante identificar el mecanismo molecular y la vía biológica subyacente a la alteración celular y la toxicidad del NP en las células y tejidos diana. Estudios in vitro recientes demostraron que el perfil de expresión génica combinado con respuestas celulares y bioquímicas ha proporcionado una evaluación directa de la perturbación celular y la toxicidad potencial de NP [7,8,9,10].
Las nanopartículas de oro (AuNP) se han utilizado como vehículo de entrega para la entrega de restos de silenciamiento génico en objetivos específicos, solos o en combinación con otros fármacos debido a sus propiedades fisicoquímicas y químicas de superficie únicas [11, 12]. La interacción de AuNP con las proteínas del plasma sanguíneo forma una corona de proteínas, que a su vez altera la química de la superficie de las NP e influye en las respuestas biológicas posteriores, como la captación celular y la toxicidad potencial [13, 14]. La captación celular de AuNP en diferentes líneas celulares de cáncer humano y células primarias se vio críticamente afectada por la formación de corona de proteínas, independientemente del tamaño y la carga superficial [7, 8, 9, 14, 15, 16, 17].
El estrés oxidativo dependiente del tamaño y de la carga superficial también se observó en la línea celular de cáncer de mama humano, MDA-MB-231, carcinoma hepatocelular HepG2 y células de leucemia humana HL-60 en respuesta a AuNP, que se asociaron con citotoxicidad de NP [18 , 19]. La citotoxicidad inducida por AuNP se produjo en varias líneas celulares de cáncer humano y células humanas primarias de una manera específica del tipo de célula [7, 8, 9, 20, 21].
Las enzimas del citocromo P450 (CYP) desempeñan un papel importante en la bioactivación o inactivación de numerosos fármacos citotóxicos y en la susceptibilidad del huésped a la carcinogenicidad de los fármacos contra el cáncer [22]. AuNP influyó en la actividad catalítica de las enzimas CYP a nivel celular y molecular in vivo e in vitro [7, 23, 24, 25]. AuNP ha exhibido considerablemente la expresión génica diferencial predominantemente involucrada en marcadores de estrés oxidativo en la línea celular de fibroblastos de pulmón humano MRC-5, y disfunción mitocondrial en células de vena umbilical humana (HUVEC) y hepatocitos humanos, que se correlaciona con un aumento en la producción de peróxido de lípidos y una alta citotoxicidad [8, 9, 26]. Si bien este conocimiento sugiere recíprocamente que AuNP causa muerte celular apoptótica o necrótica en varios tipos de células y altera las funciones celulares y bioquímicas combinadas con la expresión génica diferencial en las vías de respuesta al estrés y la toxicidad, las vías específicas a través de las cuales AuNP ejerce sus efectos tóxicos dentro de la célula o biológicos el sistema sigue siendo desconocido.
En este documento, este estudio investigó los efectos de la corona de proteínas, el tamaño y la carga superficial en la interacción de AuNP con la célula C3A de HCC humana. Principalmente, la captación celular dependiente del tiempo de AuNP de 40 y 80 nm funcionalizada con BPEI catiónico, ácido lipoico aniónico (LA) o polietilenglicol neutro (PEG) en células C3A se determinó con y sin corona de proteína plasmática humana (PC). En segundo lugar, se monitorizó la citotoxicidad inducida por AuNP y la producción de especies reactivas de oxígeno (ROS) / especies reactivas de nitrógeno (RNS) junto con sus efectos inhibidores sobre la actividad de CYP3A4. Por último, el mecanismo de acción molecular asociado a la toxicidad de AuNP se caracterizó utilizando el Buscador de vías de toxicología molecular humana y los Transportadores de fármacos humanos RT 2 Matriz de PCR Profiler ™.
Métodos
Síntesis de nanopartículas de oro
El BPEI catiónico de 40 y 80 nm, el LA aniónico y el PEG Biopure ™ AuNP neutro se sintetizaron de forma personalizada a partir de nanoComposix (San Diego, CA). El tamaño de partícula, el índice de polidispersidad (PDI) y el potencial zeta (z) y las propiedades espectrales se caracterizaron con dispersión dinámica de luz (DLS), microscopía electrónica de transmisión (TEM) y espectroscopía UV-Vis. Las AuNP se sintetizaron mediante la reducción de hidrato de tetracloroaurato (III) hidrato en solución acuosa de carbonato de potasio seguido del proceso de envejecimiento y filtración de flujo tangencial (TFF). La superficie de AuNP se funcionalizó con LA o PEG añadiendo ácido dihidrolipoico (0,2:1, w / w ) o PEG terminado en tiol-metoxi (Laysan Bio Inc., Arab, AL) (0,5:1, w / w ), respectivamente, seguido de lavado con TFF y filtración estéril. Las superficies funcionalizadas con BPEI de AuNP se sintetizaron mediante la química de EDC uniendo el ácido carboxílico de LA con aminas libres de BPEI seguido de lavado con TFF y posterior centrifugación para eliminar el BPEI no unido.
Preparación de corona de proteínas
Plasma sanguíneo humano combinado ( n =5) se obtuvieron de Biological Specialty Corp. (Colmar, PA). Las AuNP se incubaron en plasma humano (55%, v / v ) a una velocidad constante de 250 rpm a 37 ° C durante 1 h como se informa [7, 8]. Las proteínas no unidas y débilmente asociadas se eliminaron mediante lavados repetidos con solución salina tamponada con fosfato (PBS) a 20.000 × g durante 20 min a 20 ° C. El AuNP final recubierto con corona de proteína de plasma humano (PC) se dispersó en PBS y luego se diluyó en medio de cultivo celular para una caracterización o dosificación fisicoquímica adicional. El protocolo detallado se proporciona en el archivo adicional 1.
Caracterización fisicoquímica de AuNP
Diámetros hidrodinámicos ( D H ), PDI y potencial z del AuNP desnudo (sin PC) de 40 y 80 nm funcionalizado con BPEI, LA y PEG en agua desionizada (DI) se analizaron a 25 ° C a 0 h utilizando el Zetasizer Nano ZS (Malvern Instruments, Worcestershire, Reino Unido); para AuNP revestido con PC en PBS a 25 ° C a 0 h; y para todos los AuNP desnudos y PC en medio de cultivo celular completo a 37 ° C a 0 hy 24 h. El medio de cultivo celular completo contenía medio esencial mínimo de Eagle (EMEM) suplementado con FBS al 10% (ATCC ® , Manassas, VA). Se midió una muestra 5 veces con 11 sub-corridas de 10 s cada una. Además, los espectros de absorción óptica se midieron utilizando el lector de microplacas multimodo híbrido Synergy H1 (BioTek Instruments Inc., Winooski, VT) a temperatura ambiente a las 0 h.
Microscopio electrónico de transmisión
La morfología de AuNP se caracterizó mediante TEM. Toda la solución de AuNP desnuda y PC (5 μl) se colocó en rejillas de cobre de malla 200 seguido de secado al aire a temperatura ambiente. Las muestras se vieron en un Tecnai G2 Spirit BioTWIN con un detector Oxford (FEI Company, Hillsboro, OR) a un voltaje de aceleración de 120 kV. La suite de microscopía GATAN (GATAN Inc., Pleasanton, CA) midió los diámetros de AuNP.
Medición de viabilidad y cultivo celular
Células C3A de carcinoma hepatocelular humano (ATCC ® CRL-10741 ™) se adquirieron de ATCC ® (Manassas, VA), cultivado en EMEM completo (ATCC ® , Manassas, VA) suplementado con FBS al 10%, y se expandió hasta aproximadamente un 80% de confluencia en un matraz T75 con cambios de medio cada 4 días. Después del 0,25% ( w / v ) digestión con tripsina - 0,53 mM de ácido etilendiaminotetraacético (EDTA), las células se sembraron en placas de 96 pocillos a 8 × 10 4 células por pocillo y se incuban a 37 ° C en una atmósfera humidificada de 95% de aire y 5% de CO 2. Después de 48 h de incubación, las células se dosificaron con AuNP en ausencia y presencia de PC. Las células C3A entre los pases 9 y 12 se utilizaron para la dosificación.
La viabilidad de C3A se determinó utilizando alamarBlue ® ensayo de viabilidad (Thermo Sci., Waltham, MA) como se describe [7, 27]. Las células en las placas de 96 pocillos se trataron con BPEI-, LA- y PEG-AuNP de 40 y 80 nm con y sin PC en un rango de 0 a 250 μg / cm 2 . Después de 24 h, 10% de alamarBlue ® reactivo en EMEM completo ( v / v ) al cultivo celular y se incubó durante 3 ha 37 ° C. El EMEM completo sirvió como dispersante. Las interacciones de AuNP con el ingrediente activo de alamarBlue ® reactivo, resazurina o un producto reducido, resorufina se midieron como controles. AuNP y resazurina (sin células) o medio de mantenimiento (sin células) sirvieron como controles de fondo. La fluorescencia, proporcional a la viabilidad celular, se normalizó a los controles y se expresó como un porcentaje relativo al grupo de células de control.
Medición de la absorción celular con espectrometría de masas de plasma acoplado inductivamente
Las células se sembraron a 8 × 10 4 células por pocillo de placas de 96 pocillos y dosificadas con una concentración no tóxica de 1,56 μg / cm 2 de todos los AuNP desnudos y PC durante 0,5, 1, 3, 6, 12 y 24 h. El paso de grabado se incorporó para eliminar el AuNP unido a la membrana celular y su unión no específica a los pozos, como se informó anteriormente [28]. La cosecha de células se secó y digirió en agua regia y la concentración de Au intracelular se cuantificó utilizando el NexION ™ Espectrometría de masas de plasma acoplado inductivamente 350X (ICP-MS) (PerkinElmer, Waltham, MA). La captación celular de AuNP se calculó como se informó anteriormente y se expresó como el número de AuNP por célula [29]. El protocolo detallado se proporciona en el archivo adicional 1.
Mediciones de estrés oxidativo / nitrosativo
Las células se sembraron a 8 × 10 4 células por pocillo de placas de 96 pocillos y dosificadas con BPEI y PEG-AuNP desnudos de 40 nm hasta 125 μg / cm 2 durante 1, 3 y 24 h. La medición directa de oxígeno / estrés nitrosativo se ensayó con el kit de ensayo de especies reactivas de oxígeno total (ROS) / superóxido (SO) (Enzo Life Sciences, Farmingdale, NY) como se describió anteriormente [30]. Se midió la fluorescencia, proporcional al aumento de ROS / especies de nitrógeno reactivo (RNS) (Ex488 / Em520 nm) o SO (Ex550 / Em610 nm) con el lector de microplacas. El protocolo detallado se proporciona en el archivo adicional 1.
Actividad del citocromo P450 3A4
Los efectos adversos de 40 y 80 nm desnudos y PC AuNP sobre la actividad CYP3A4 se caracterizaron utilizando ensayos P450-Glo ™ (Promega Corp., Madison, WI) como se describe en su totalidad [7]. Se dosificaron células C3A en placas de 96 pocillos a la concentración letal media (LC 50 ) valores:127,3 μg / cm 2 del BPEI-AuNP de 40 nm, 205,5 μg / cm 2 del BPEI-AuNP de 80 nm, 192,5 μg / cm 2 del LA-AuNP de 40 nm y 129,5 μg / cm 2 del PEG-AuNP de 40 nm. Desde LC 50 No se determinaron los valores de LA- y PEG-AuNP de 80 nm, las células se trataron con LC 50 valores de 40 nm LA- y PEG-AuNP (192,5 μg / cm 2 y 129,5 μg / cm 2 , respectivamente). Después del final de la incubación de 24 h, las células se incubaron con un sustrato de CYP3A4 (luciferina-IPA) a 37 ° C durante 3 h. La señal de luminiscencia, proporcional a la actividad enzimática, se midió con un lector de microplacas y luego se normalizó a los controles. Se asignaron controles para evaluar la interacción de AuNP con sustratos o metabolitos parentales y sustratos libres de células. La actividad CYP se expresó como un porcentaje relativo al grupo de células de control.
Perfiles de expresión genética
Dado que se emplearon PEG-AuNP tóxicos de 40 nm en la inhibición de la actividad CYP3A4 y la actividad antioxidante en células C3A que muestran una alta captación celular, se seleccionó para caracterizar los mecanismos moleculares de acción subyacentes a su toxicidad y respuestas celulares diferenciales. Las células se sembraron a 2,5 × 10 6 células por pocillo de placas de 6 pocillos y dosificadas con LC 50 valor del PEG-AuNP de 40 nm durante 24 ha 37 ° C. Al final de la incubación, las células se sometieron a aislamiento de ARN y luego se realizó la síntesis de ADNc utilizando ARN total con un valor promedio de integridad de ARN (RIN) de 7.8 como se describió anteriormente [7, 8, 9]. El ADNc resultante se mezcló con RT 2 SYBR green master mix (Qiagen Inc., Valencia, CA) y luego se aplicó al Buscador de vías de toxicología molecular humana o Transportadores de fármacos humanos RT 2 Matrices de PCR Profiler ™ en Quantstudio ™ 7 Flex (Applied BioSystem, Foster City, CA). Genes expresados diferencialmente con el cambio de veces <- 2 y> 2 y una p <0,05 representaba regulación ascendente y descendente del gen de interés. Para validar el RT 2 Los datos de la matriz de PCR, se evaluó una expresión de nueve genes seleccionados con síntesis de ADNc y posterior PCR en tiempo real. Las secuencias de cebadores se resumen en el archivo adicional 1:Tabla S1. Todas las reacciones de PCR se realizaron por triplicado. El protocolo detallado de las condiciones de PCR en tiempo real y la cuantificación se proporciona en el archivo adicional 1.
Análisis estadístico
Concentración letal media (LC 50 ) Los valores de AuNP en células C3A se estimaron ajustando una ecuación de Hill con pendiente variable a los datos observados (la entrada de los niveles de concentración de AuNP y la viabilidad celular correspondiente) utilizando GraphPad Prism 6 (La Jolla, CA) como se describe [7]. Se realizó un análisis de varianza unidireccional (ANOVA) utilizando SAS 9.4 (SAS Institute, Cary, NC) para evaluar los efectos de AuNP en la producción de ROS / RNS y la captación celular en las células C3A. Si fue significativa, la comparación múltiple se realizó con la prueba de diferencia significativa honesta (HSD) de Tukey en una p <0.05.
Resultados y discusión
Caracterización fisicoquímica de PC AuNP de plasma desnudo y humano
Los efectos del tamaño de NP, la carga superficial y la formación de PC en plasma humano alrededor de AuNP sobre el diámetro hidrodinámico (D H ), el índice de polidispersidad (PDI), el potencial z y una propiedad espectral, así como la morfología, se han caracterizado mediante espectroscopía DLS, TEM y UV-Vis (Fig. 1). En las imágenes TEM, todo el AuNP desnudo (sin PC) en agua DI estaba monodisperso con la distribución de tamaño ajustada y los rangos de espectro UV-Vis únicos de 521–553 nm (Fig. 1a). Se observaron formaciones de PC alrededor de AuNP con los cambios en la distribución del tamaño y los desplazamientos al rojo de los espectros de absorción (Fig. 1b). La D H y los valores de PDI de 40 y 80 nm desnudo y PC AuNP en EMEM completo fueron compatibles hasta 24 ha 37 ° C excepto para el PC BPEI-AuNP de 40 y 80 nm que mostraron una disminución en los valores de PDI (0.29 y 0.32, respectivamente ) a las 24 ha 37 ° C en comparación con las de 0 ha 37 ° C (0,62 y 1,0, respectivamente) (Tabla 1). Los valores de potencial Z de todos los AuNP desnudos y PC disminuyeron relativamente a las 24 ha 37 ° C en comparación con los de 0 ha 37 ° C. Se observó una agregación de PC BPEI-AuNP de 40 y 80 nm en PBS y EMEM completo, que se correlacionó con múltiples picos en una distribución de tamaño y cambios en D H y corrimientos al rojo de los espectros de absorbancia en relación con el BPEI-AuNP desnudo (Fig. 1 y archivo adicional 1:Figura S1, Tabla 1). Estos resultados fueron respaldados por estudios recientes de que el PC de 40 y 80 nm y el BPEI-AuNP recubierto por corona de albúmina de suero humano se agregaron en PBS y varios medios de cultivo celular [7, 8, 9].
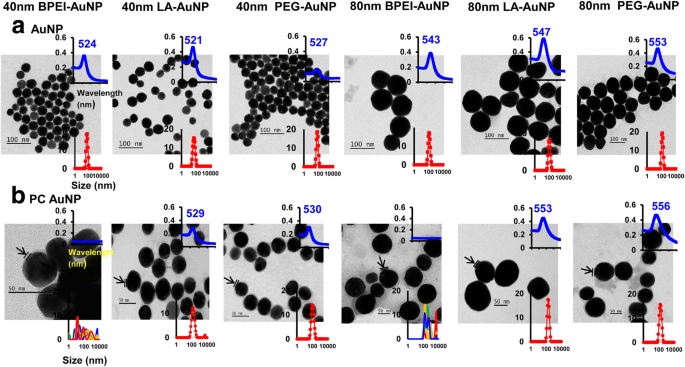
Micrografías electrónicas de transmisión de a AuNP en agua desionizada y b PC AuNP en PBS a 0 ha 25 ° C, longitud de onda de los espectros UV-Vis (recuadro superior) y distribución dinámica de la dispersión de luz (recuadro inferior). Las flechas indican la formación de PC. PC corona de proteínas plasmáticas humanas, BPEI polietilenimina ramificada, LA ácido lipoico, PEG polietilenglicol
Citotoxicidad AuNP
La citotoxicidad de AuNP se midió utilizando la concentración letal media (LC 50 ) en células C3A. La carga superficial de NP, el tamaño de partícula y la formación de PC alrededor de LC 50 dependiente de NP Los análisis con AuNP se muestran en la Fig. 2. Todas las BPEI-, LA y PEG-AuNP de 40 nm y las BPEI-AuNP de 80 nm fueron citotóxicas para las células C3A con la LC 50 correspondiente oscila entre 127,3 y 205,5 μg / cm 2 (Figura 2a). El PEG-AuNP desnudo de 80 nm mostró una viabilidad celular del 59% a la concentración más alta de 250 μg / cm 2 , mientras que los LA-AuNP de 80 nm no fueron citotóxicos. La PC redujo la toxicidad de AuNP en función del tamaño y la modificación de la carga superficial, excepto por el BPEI-AuNP de 80 nm que mostró una viabilidad celular del 51% a 250 μg / cm 2 a las 24 h (Fig. 2b). Estudios recientes demostraron que el BPEI-AuNP desnudo de 40 nm era tóxico para los hepatocitos humanos primarios, HUVEC y las células del túbulo proximal renal humano (HPTC) (LC 50 rangos de 22,4 a 80,3 μg / cm 2 ) [7,8,9]. Los BPEI-AuNP recubiertos con PC eran citotóxicos para los hepatocitos humanos, pero los AuNP recubiertos con HSA no eran citotóxicos [7]. Estos resultados sugirieron que las células C3A son más resistentes a la toxicidad de AuNP que las células humanas primarias debido a una alta tasa de proliferación y actividad metabólica de la línea celular cancerosa [31].
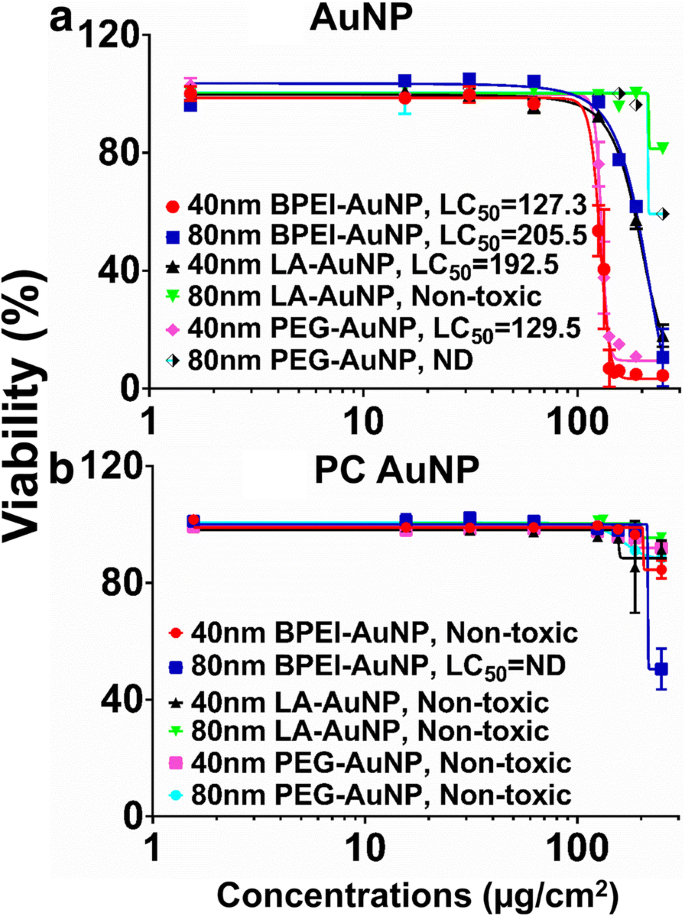
Viabilidad de C3A y LC 50 valores de 40 y 80 nm a AuNP y b PC AuNP. Los datos representan la media ± S.D. ( n =3). Corona de proteína plasmática humana PC, ND no determinado, polietilenimina ramificada BPEI, ácido lipoico LA, polietilenglicol PEG, LC 50 concentración letal media
Captación intracelular de AuNP
La captación celular dependiente del tamaño de NP, de la carga superficial y de la PC de todas las AuNP desnudas y de PC se determinó en 1,56 μg / cm 2 hasta 24 h. ANOVA mostró cambios significativos con el tamaño, el PC y el tiempo ( p <0,0001) e interacciones (PC × tamaño, PC × tiempo, tamaño × tiempo y PC × tamaño × tiempo) ( p <0,001) para toda la captación de AuNP además de una interacción insignificante (PC × tamaño) para la captación de LA- y PEG-AuNP ( p =0,2). Como se muestra en la Fig. 3a-f, se observó un aumento lineal en la captación celular de 40 y 80 nm desnudo y PC AuNP además de 40 nm desnudo y PC BPEI-AuNP que alcanzaron la mayor captación celular a las 6 h y luego disminuyó ( Figura 3a). Sin embargo, a las 24 h, el BPEI-AuNP catiónico de 40 nm contenía la absorción más alta seguida de PEG-AuNP neutro de 40 nm y luego LA-AuNP aniónica de 40 nm, que se asoció con el orden de citotoxicidad de las células C3A de AuNP (Fig. 2a ). Este resultado es consistente con los estudios previos de que la poli (N- (2-aminoetil) acrilamida) catiónica y BPEI-AuNP tuvieron la mayor absorción celular en comparación con las del poli (ácido acrílico) aniónico, y LA-AuNP y poli neutro ( N- (2,3-dihidroxipropil) acrilamida y PEG-AuNP en células Caco-2 de adenocarcinoma colorrectal humano, HPTC y hepatocitos humanos [9, 32]. Además, el complejo NP-PC atenuó todos los BPEI de 40 y 80 nm. y LA-AuNP y el PEG-AuNP de 40 nm en las células C3A, pero aceleraron la captación de PEG-AuNP de 80 nm (Fig. 3f). Estos resultados están respaldados por estudios recientes de que la PC inhibe la captación de AuNP en HUVEC, HEK y HPTC, independientemente del tamaño y la carga superficial [8, 9, 33]. Por el contrario, las coronas de PC y HSA mejoraron la captación de PEG-AuNP de 40 nm en los hepatocitos humanos, pero esta última indujo la captación de PEG-AuNP de 80 nm en HEK [7, 33] .
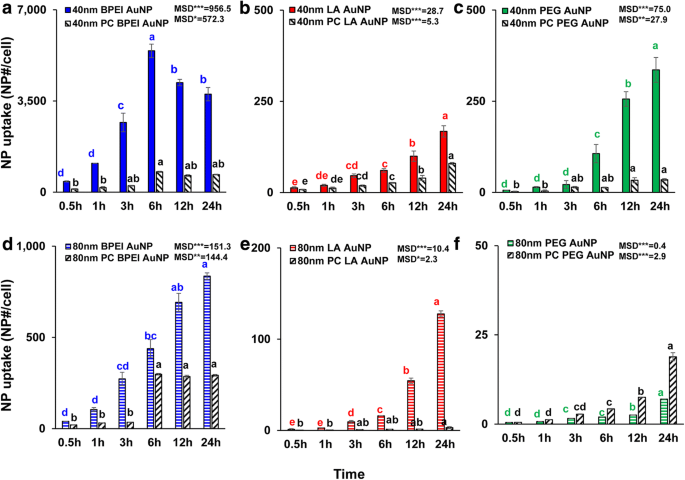
Captación celular dependiente del tiempo de 40 nm a BPEI-AuNP, b LA-AuNP y c PEG-AuNP y el d de 80 nm BPEI-AuNP, e LA-AuNP y f PEG-AuNP en ausencia y presencia de PC en células C3A hasta 24 h. Los datos representan la media ± S.D. ( n =3). Las letras fueron significativamente diferentes según la prueba HSD de Tukey. BPEI polietilenimina ramificada, LA ácido lipoico, PEG polietilenglicol, PC corona de proteína plasmática humana, MSD diferencia mínima significativa. * p <0,05; ** p <0,005; *** p <0,0001
Medidas de estrés oxidativo y nitrosativo
Dado que el BPEI y el PEG-AuNP desnudos de 40 nm exhibieron una mayor citotoxicidad y absorción celular en las células C3A en comparación con otros AuNP, se seleccionaron para investigar el estrés oxidativo / nitrosativo inducido por AuNP. Ambos AuNP modularon la generación de ROS / RNS en células C3A de manera dependiente del tiempo y de la concentración ( p <0,0001) y por interacción (tiempo × concentración, p <0,0001). Como se muestra en la Fig. 4a, la generación de ROS / RNS disminuyó a las concentraciones más altas de BPEI-AuNP de 40 nm (62,5 μg / cm 2 y 125 μg / cm 2 ) a 1 ha 37 ° C pero aumentó hasta 24 h. Por el contrario, el PEG-AuNP de 40 nm suprimió sustancialmente la generación de ROS / RNS a 39,1 μg / cm 2 en adelante con un cambio de pliegue <0,5 hasta 24 h (Fig. 4b). La activación de la muerte celular a menudo contribuye a la toxicidad del NP y, en la mayoría de los casos, un aumento en la producción de ROS / RNS, que conduce al estrés oxidativo, es responsable de la toxicidad del NP [34]. La producción de ROS / RNS dependiente de la carga superficial se observó con BPEI catiónico de 40 nm y PEG-AuNP neutro. El BPEI-AuNP de 40 nm mostró un patrón bifásico de generación de ROS / RNS (antioxidante a la 1 hy prooxidante a las 3 h en adelante) a altas concentraciones, que se asoció con su citotoxicidad en las células C3A (Fig. 2a). Este resultado es consistente con los estudios previos de que la generación de ERO inducida por BPEI-AuNP de 40 y 80 nm y la generación de ROS inducida por citrato-AuNP de 20 nm se asoció con sus citotoxicidades en hepatocitos humanos y células HepG2, respectivamente, de manera dependiente del tiempo y de la concentración. [7, 35]. AuNP mostró citotoxicidad inducida por estrés oxidativo en células de leucemia promielocítica humana, HL-60 con una reducción total de glutatión, independientemente del tamaño [19]. Por el contrario, el PEG-AuNP de 40 nm sirvió como antioxidante, lo que sugiere que el estrés oxidativo / nitrosativo puede no ser un mecanismo directo de la citotoxicidad inducida por PEG-AuNP de 40 nm en las células C3A (Fig. 2b).
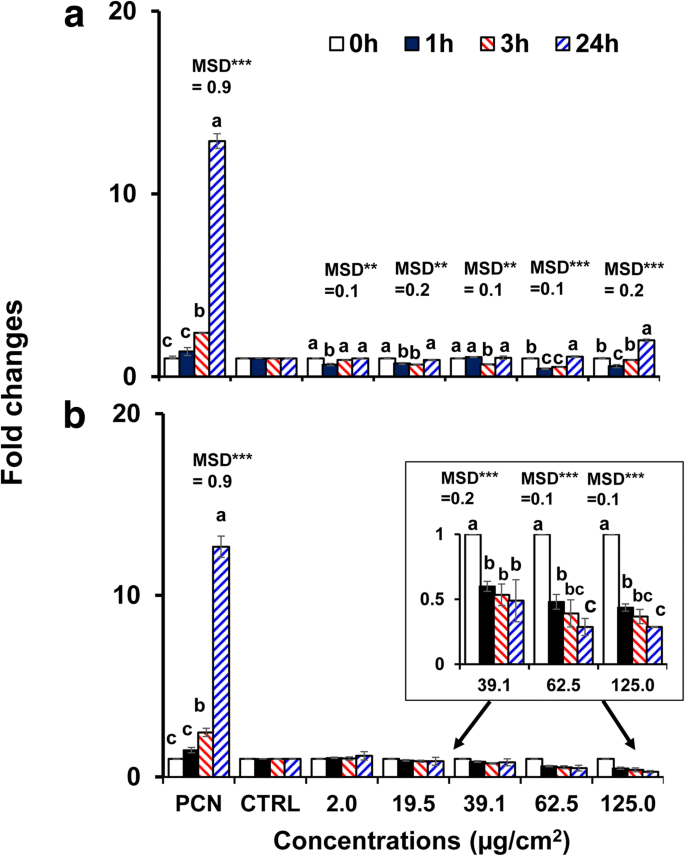
Producción de ROS / RNS dependiente del tiempo y de la concentración en células C3A expuestas a a el BPEI-AuNP de 40 nm y b el PEG-AuNP de 40 nm hasta 24 h. Los datos representan la media ± S.D. ( n =3). Las letras fueron significativamente diferentes según la prueba HSD de Tukey. BPEI polietilenimina ramificada, LA ácido lipoico, PEG polietilenglicol, CTRL control, MSD una diferencia mínima significativa, PCN piocianina (inductor de ROS). ** p <0,005; *** p <0,0001
Medición de la actividad CYP3A4
Se caracterizaron los efectos inhibidores de 40 y 80 nm desnudos y PC AuNP sobre la actividad de CYP3A4. Como se muestra en la Fig.5, el BPEI-, LA- y PEG-AuNP de 40 nm y el BPEI-AuNP de 80 nm en LC 50 Los valores inhibieron la actividad catalítica de CYP3A4 en células C3A con la correspondiente actividad de 20,1 a 31,4% en relación con los controles, independientemente del tamaño y la carga superficial. Las concentraciones no tóxicas de LA- y PEG-AuNP de 80 nm también suprimieron su actividad (31,4 y 26,6%, respectivamente). Sin embargo, la PC mejoró ampliamente la inhibición de CYP3A4 inducida por AuNP de 40 y 80 nm además de la PEG-AuNP de 40 y 80 nm que muestra una actividad del 63 y 46% en comparación con los controles. Esto concuerda con estudios in vitro con tejido hepático humano y hepatocitos en los que el ácido tánico aniónico-AuNP y el BPEI-AuNP catiónico de 40 y 80 nm inhibieron sustancialmente la actividad catalítica de CYP3A4 [7, 25]. En contraste, PEI-AuNP catiónico y polivinilpirrolidona-AuNP neutra inducen la expresión de ARNm de CYP1A2, CYP2C9 y CYP3A4 en células HepG2 y CYP2B y CYP3A en corte de hígado de rata, respectivamente [36, 37]. Un estudio reciente informa que el BPEI-AuNP desnudo y PC de 40 y 80 nm suprimió sustancialmente la actividad de CYP3A4 en los hepatocitos humanos mediante un cambio conformacional en la proteína o bloqueando el bolsillo del sustrato como una inhibición reversible [7].
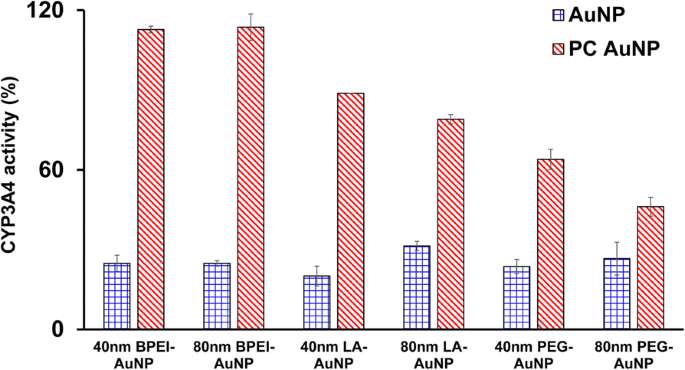
Un efecto inhibidor de AuNP sobre la actividad de CYP3A4 en células C3A expuestas a BPEI-, LA- y PEG-AuNP de 40 y 80 nm en ausencia y presencia de PC durante 24 h. Los valores representan la media ± S.D. ( n =3). BPEI polietilenimina ramificada, LA ácido lipoico, PEG polietilenglicol, PC corona de proteínas plasmáticas humanas
Perfil de expresión génica centrada en la vía de toxicidad del PEG-AuNP de 40 nm
De los genes representativos que cubren 13 vías diferentes de estrés y toxicidad, se expresó diferencialmente un total de 212 genes (↓ 186 y ↑ 26 genes) en LC 50 valor del PEG-AuNP de 40 nm (Fig. 6, archivo adicional 1:Tablas S2-S7). El 12,3% (26 genes, ↓ 26, ↑ 0 genes) del total de genes (212 genes) participaron predominantemente en la β-oxidación de ácidos grasos mitocondriales; para apoptosis 11,3% (24 genes, ↓ 18, ↑ 6 genes); para la vía de reparación y daño del ADN 11,3% (24 genes, ↓ 18, ↑ 6 genes); y para la respuesta al choque térmico 11,3% (24 genes, ↓ 22, ↑ 2).
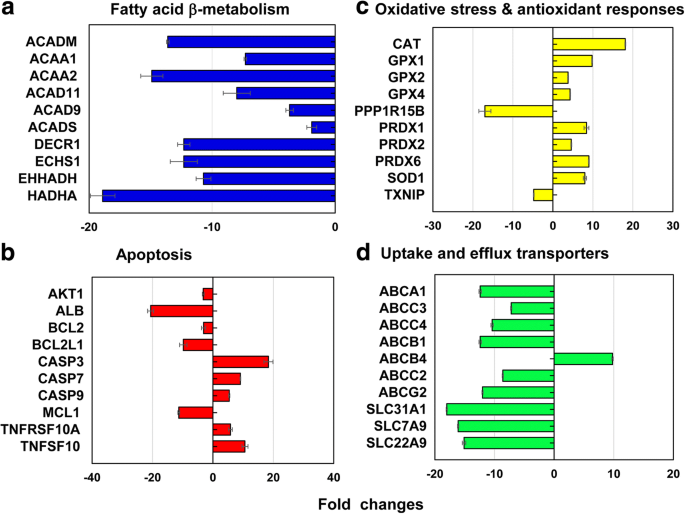
Genes representativos implicados en a β-oxidación de ácidos grasos mitocondriales, b apoptosis, c estrés oxidativo y respuestas antioxidantes, y d transportadores de captación y eflujo hepático a LC 50 valor de 40 nm de PEG-AuNP. Todos los datos tuvieron un cambio de veces <- 2 y> 2 en p <0,05. El análisis de ontología genética se enumera en el archivo adicional 1:Tablas S2 – S7
En la vía de oxidación β de ácidos grasos mitocondriales, genes que codifican tres enzimas diferentes involucradas en la producción de acil-CoA y equivalentes reductores de NADH y FADH 2 fueron principalmente suprimidos; Genes ACAD11, ACAD9, ACADM y ACADS en acil-CoA deshidrogenasas (2,0 a 13,6 veces); ACAA1 y ACAA2 en cetoacil-CoA tiolasas (7,3 a 14,9 veces); DECR1, ECHS1, EHHADH y HADHA (10,7 a 18,9 veces) en enoil-CoA hidratasa (Fig. 6a, archivo adicional 1:Tabla S2). La β-oxidación de ácidos grasos mitocondriales juega un papel importante en la producción de acil-CoA y en la reducción de equivalentes de NADH y FADH 2 , que se asocia con cuatro enzimas principales (acil-CoA deshidrogenasas, enoil-CoA hidratasas, hidroxiacil-CoA deshidrogenasas y cetoacil-CoA tiolasas [38, 39]). Además, los portadores de electrones, NADH y FADH 2 , están involucrados en el ciclo del ácido tricarboxílico (TCA) y la cadena respiratoria mitocondrial, lo que resulta en la producción de ATP. En el estudio actual, el PEG de 40 nm indujo una disfunción mitocondrial, una pérdida de mantenimiento de ATP a través de una disminución en los niveles intracelulares de ATP y FADH 2 , definiendo consecuentemente su citotoxicidad en células C3 (Fig. 2a). El fenómeno similar se informó en hepatocitos humanos, HUVEC y HPTC, expuestos a 40 nm BPEI-AuNP, lo que indica que la disfunción mitocondrial puede ser un mecanismo común de toxicidad AuNP, independientemente de la carga superficial y los tipos de células [7,8,9]. Un estudio reciente informó que se observó citotoxicidad relevante para la disfunción mitocondrial en células epiteliales de cáncer de próstata inmortalizadas y células epiteliales de cáncer de pulmón en respuesta a un inhibidor de la fosforilación de STAT3, OPB-51602 [40].
En la vía de la apoptosis, los seis genes proapoptóticos de CASP3, CASP7, CASP9, TNFRSF10A, TNFRSF10B y TNFSF10 estaban regulados positivamente, mientras que los seis genes antiapoptóticos de AKT1, ALB, BCL2, BCL2L1, MCL1 y XIAP estaban regulados negativamente (Fig. .6b, archivo adicional 1:Tabla S3), que se correlacionó con la citotoxicidad dependiente de la dosis en las células C3A (Fig. 2a). En el punto de control de reparación y daño del ADN, los genes de las quinasas del punto de control (CHEK1 / 2), los genes de reparación por escisión del ADN (ERCC1 / 2/3) y la ligasa IV del ADN (LIG4) se regularon positivamente, pero otros genes de reparación por escisión (ERCC5 / 6, XRCC1 / 5), los genes de la quinasa del punto de control (CDKN1A) y de las proteínas quinasas (PRKDC) se regularon negativamente (de 2 a 19 veces). Estos resultados sugirieron que la interferencia inducida por PEG-AuNP de 40 nm con el ciclo celular y el sistema de reparación del ADN puede correlacionarse con una inducción de la muerte celular en las células C3A (Fig. 2a, archivo adicional 1:Tabla S3). Los genes que codifican dos proteínas de choque térmico (HSP) diferentes (A1A y A1B) se regularon al alza (10,2 a 14,2 veces) pero HSP40 subfamilia A, B y C; Miembro 1 de HSP90; y HSP60 fueron regulados a la baja (de 2 a 16 veces) (Archivo adicional 1:Tabla S4).
En el estrés oxidativo y la respuesta antioxidante, el PEG-AuNP de 40 nm en LC 50 valoran los genes antioxidantes inducidos y los prooxidantes suprimidos, lo que se asoció con una disminución en la generación de ROS / RNS siendo antioxidante en sí mismo (Fig. 4b). En los genes antioxidantes, se indujo glutatión peroxidasa (GPX) 1, GPX2, GPX4, PRDX1, PRDX2, PRDX6, superóxido dismutasa (SOD) 1 y CAT (3,8 a 18,1 veces). En los genes prooxidantes, se suprimieron TXNIP y PPP1R15B (4,8 y 17 veces, respectivamente) (Fig. 6c, archivo adicional 1:Tabla S5). Esto es consistente con un estudio previo que AuNP mostró citotoxicidad inducida por estrés oxidativo en HepG2 y hepatocitos humanos, independientemente del tamaño [7, 19].
En el metabolismo de fase I, los genes CYP3A4 y ESD se suprimieron en gran medida (7 y 12 veces, respectivamente). Especialmente, el efecto inhibidor de PEG-AuNP de 40 nm sobre la expresión de CYP3A4 se correlacionó con una disminución en la actividad de CYP3A4 (Fig. 5). Estudios recientes informaron que el BPEI-AuNP de 40 nm inhibía la expresión génica de CYP1A2, CYP2C9 y CYP3A4 en hepatocitos humanos; ESD en HUVEC; y CYP1A1 en HPTC [7, 8, 9]. El estudio epidemiológico demostró que las enzimas CYP en el tejido hepático de pacientes con CHC fueron inhibidas sustancialmente por el proceso tumorigénico a nivel molecular y funcional [41].
Perfil de expresión génica del transportador de eflujo y absorción de fármacos
El desarrollo de resistencia a múltiples fármacos (MDR) por parte de las células tumorales es una de las principales causas de los fracasos del tratamiento del cáncer [42, 43]. La disminución de la absorción del fármaco mediada por transportadores de membrana integrales y el aumento del flujo de salida del fármaco, incluida la glicoproteína P (P-gp) y la proteína de resistencia al cáncer de mama (BCRP), es uno de los principales mecanismos de la MDR.
La expresión génica diferencial de los transportadores de captación y salida de fármacos en células C3A expuestas a PEG-AuNP de 40 nm mostró que un total de 14 genes de transportadores ABC (↓ 12 y ↑ 2 genes) y un total de 21 genes de transportadores SLC (↓ 21 y ↑ 0 genes) se modularon sustancialmente en LC 50 valor (Figs. 6d y 7, archivo adicional 1:Tabla S7). En los transportadores de eflujo de fármacos de la familia ABC, los genes de la proteína asociada a la resistencia a múltiples fármacos (MRP3 / ABCC3), MRP4 (ABCC4) y la proteína regulada por el flujo de salida de colesterol (CERP / ABCA1) en la membrana basolateral se regularon negativamente (7,2 a 10,4 veces). Los genes que codifican P-gp (ABCB1), MRP2 (ABCC2), BCRP (ABCG2) y esterolina 2 (ABCG8) en los transportadores de flujo de salida canalicular también se suprimieron (8,6 a 13,8 veces). Por el contrario, la resistencia a múltiples fármacos (MDR4 / ABCB4) en la membrana canalicular y el transportador ABC de las mitocondrias (MTABC3 / ABCB6) en la membrana mitocondrial externa estaban altamente reguladas al alza (9,8 y 5,8 veces, respectivamente). En los transportadores de absorción de fármacos, también se inhibieron los genes de la proteína transportadora de cobre (CTR1 / SLC31A1) y, en menor grado, el transporte de aniones orgánicos (OAT7 / SLC22A9) (18 y 15 veces, respectivamente). Estos resultados apoyan un estudio reciente de que el BPEI-AuNP de 40 nm regula a la baja MDR3 en hepatocitos humanos pero regula al alza MRP3 en HUVEC, lo que indica una interacción dependiente de la carga superficial y del tipo celular entre AuNP y transportadores de salida [7, 8]. El estudio epidemiológico mostró que se producía una alta expresión de BCRP y una baja expresión de OCT3 en el tumor de CHC, que estaba estrechamente asociado con la progresión del tumor y su tamaño [44]. Un estudio anterior mostró que el inhibidor de la P-gp, el verapamilo, mejoraba la citotoxicidad del glutatión-AuNP conjugado con doxorrubicina en líneas celulares de fibrosarcoma felino al aumentar la concentración intracelular del fármaco [45]. El estudio actual enfatiza que la información derivada de los mecanismos en el PEG-AuNP de 40 nm identificó una acción separada pero aún complementaria sobre la β-oxidación de los ácidos grasos mitocondriales, el ciclo del TCA y la cadena respiratoria, los transportadores de flujo y captación del fármaco, así como la actividad del CYP3A4 en Células C3A (Fig. 7). Al final, esto destacará la interacción AuNP con procesos biológicos clave y su mecanismo molecular subyacente en el HCC, que puede estar aún más implicado en el desarrollo de una diana terapéutica más eficaz en el tratamiento del HCC.
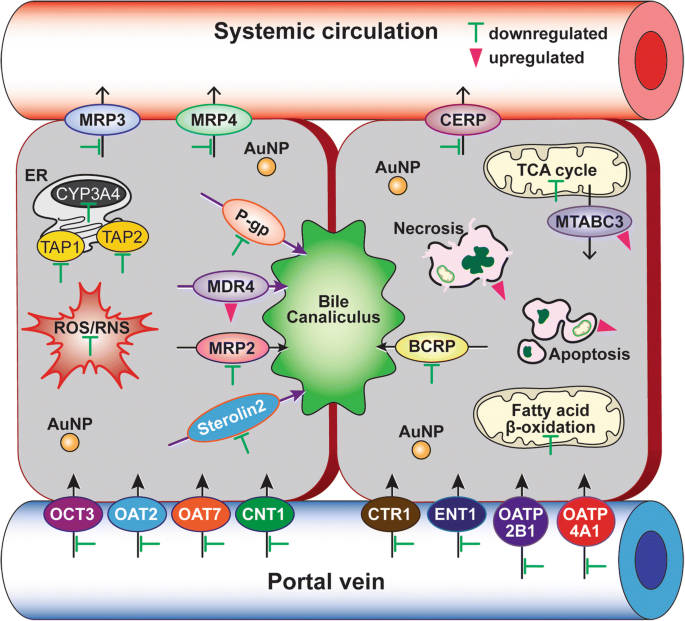
Una representación esquemática de los mecanismos básicos de acción de PEG-AuNP de 40 nm en el tratamiento del HCC. Las barras verdes (una inhibición) y los triángulos rosas (una inducción) indican las rutas y marcadores biológicos modificados con PEG-AuNP de 40 nm. El análisis de ontología genética se enumera en el archivo adicional 1:Tablas S2 – S7
Para validar el análisis de expresión génica de RT 2 matriz, los nueve genes se seleccionaron para PCR en tiempo real. En el archivo adicional 1:Tabla S1, los nueve genes se modularon en LC 50 del PEG-AuNP de 40 nm. Estos cambios transcripcionales fueron consistentes con los del análisis de expresión génica con matrices de PCR (Fig. 6, Archivo adicional 1:Tablas S2-S7).
Conclusiones
Hemos presentado que la interacción catiónica BPEI, aniónica LA o PEG-AuNP neutral con la corona de proteína plasmática humana (PC) causó los cambios en D H , PDI y potencial z de AuNP e influyeron aún más en las respuestas celulares en las células C3A. Todos los AuNP desnudos (sin PC) de 40 y 80 nm eran citotóxicos para las células C3A además del LA-AuNP de 80 nm, pero el PC mejoró completamente sus citotoxicidades además del BPEI-AuNP de 80 nm. El BPEI-AuNP desnudo de 40 nm mostró la captación celular más alta seguida por el PEG-AuNP de 40 nm y luego el LA-AuNP de 40 nm, mientras que la PC suprimió la captación de AuNP además del PEG-AuNP de 80 nm. El BPEI-AuNP de 40 nm provocó respuestas bifásicas de estrés oxidativo (pro y antioxidante) en las células C3A, mientras que el PEG-AuNP de 40 nm fue antioxidante. La actividad de CYP3A4 fue ampliamente suprimida por todas las AuNP desnudas, independientemente del tamaño y las cargas superficiales, mientras que la PC mejoró sustancialmente su efecto inhibidor sobre la actividad enzimática además de la PEG-AuNP de 40 y 80 nm. Genes expresados diferencialmente en LC 50 El valor de 40 nm de PEG-AuNP participaron principalmente en la β-oxidación de ácidos grasos mitocondriales y, en menor grado, en los transportadores de eflujo / captación hepáticos. El PEG-AuNP de 40 nm inhibió tres enzimas principales en la β-oxidación (acil-CoA deshidrogenasa, enoil-CoA hidratasa y cetoacil-CoA tiolasa), otras enzimas en el ciclo del TCA y la cadena respiratoria mitocondrial para la producción de ATP. El PEG-AuNP de 40 nm aumentó la expresión de genes proapoptóticos y disminuyó los genes antiapoptóticos en LC 50 valor. Se observó un alto nivel de antioxidantes y un bajo nivel de genes prooxidantes en células C3A expuestas a PEG-AuNP de 40 nm. Además, los genes de los transportadores de captación y salida del fármaco ubicados en la membrana basolateral y canalicular se modularon sustancialmente.
Abreviaturas
- ANOVA:
-
Análisis de varianza unidireccional
- AuNP:
-
Nanopartículas de oro desnudas:sin PC
- BPEI:
-
Polietilenimina ramificada
- CYP:
-
Citocromo P450
- D H :
-
Diámetros hidrodinámicos
- DLS:
-
Dispersión de luz dinámica
- EDTA:
-
Ácido etilendiaminotetraacético
- EMEM:
-
Medio esencial mínimo de Eagle
- HCC:
-
Carcinoma hepatocelular humano
- HPTC:
-
Células tubulares proximales renales humanas
- HSD:
-
Prueba honesta de diferencias significativas de Tukey
- HUVEC:
-
Células de la vena umbilical humana
- ICP-MS:
-
Espectrometría de masas de plasma acoplado inductivamente
- LA:
-
Ácido lipoico aniónico
- LC 50 :
-
Concentración letal media
- MDR:
-
Resistencia a múltiples fármacos
- NP:
-
Nanopartículas
- PBS:
-
Solución salina tamponada con fosfato
- PC:
-
Corona de proteínas plasmáticas humanas
- PDI:
-
Índice de polidispersidad
- PEG:
-
Polietilenglicol neutro
- ARNi:
-
Interferencia de ARN
- RNS:
-
Especies reactivas de nitrógeno
- ROS:
-
Especies reactivas de oxígeno
- Entonces:
-
Superóxido
- TEM:
-
Microscopía electrónica de transmisión
- TFF:
-
Filtración de flujo tangencial
Nanomateriales
- Mantenimiento de hoteles y su impacto en sus resultados
- Nanobiosensor de oro basado en la resonancia de plasmón superficial localizado es capaz de diagnosticar la brucelosis humana, presentando un método rápido y asequible
- Síntesis y actividad de oxidación del CO de óxido binario mixto 1D CeO2-LaO x catalizadores de oro soportados
- Síntesis de calentamiento de estado sólido de compuesto de poli (3,4-etilendioxitiofeno) / oro / grafeno y su aplicación para la determinación amperométrica de nitrito y yodato
- Promoción del crecimiento celular SH-SY5Y mediante nanopartículas de oro modificadas con 6-mercaptopurina y un péptido penetrante de neuronas
- Anticuerpo monoclonal de heparanasa marcada con nanopartículas de oro magnético y su aplicación posterior para la obtención de imágenes por resonancia magnética tumoral
- Efectos del fullereno C60 sobre la interacción del difenil-N- (tricloroacetil) amidofosfato con el ADN in silico y su actividad citotóxica contra la línea celular leucémica humana in vitro
- Evaluación de la toxicidad de las nanopartículas de PEG-PCCL e investigación preliminar sobre su efecto antitumoral de la carga de paclitaxel
- Influencia de los nanotubos de carbono y sus derivados en las células tumorales in vitro y parámetros bioquímicos, composición de la sangre celular in vivo
- Evaluación de la actividad del citocromo P450 3A4 inhibido por nanopartículas de oro y los mecanismos moleculares que subyacen a su toxicidad celular en la línea celular de carcinoma hepatocelular …
- Nanoclusores de oro de neuroproteína fluorescente:síntesis y aplicaciones en la detección de lectina vegetal y la obtención de imágenes celulares