


El microARN-133a-3p restaurado o el PSAT1 empobrecido restringen el aneurisma intracraneal inducido por el daño de las células endoteliales mediante la supresión de la vía GSK3β / β-catenina
Resumen
No está claro el papel funcional del microARN-133a-3p (miR-133a-3p) en el aneurisma intracraneal (IA). Por lo tanto, el objetivo del presente estudio fue investigar el papel regulador de miR-133a-3p en la regulación de la IA inducida por lesión endotelial vascular a través de la señalización de fosfoserina aminotransferasa 1 (PSAT1) / glucógeno sintasa quinasa 3β (GSK3β) / β-catenina ruta. Se obtuvieron tejidos normales de arteriolas intracraneales y tejidos IA de pacientes con traumatismo cerebral e IA. La expresión de miR-133a-3p, PSAT1, GSK3β y β-catenina en tejidos se determinó mediante RT-qPCR y análisis de transferencia Western. Las células endoteliales (CE) del IA humano se cultivaron y trataron con miR-133a-3p mimic y si-PSAT1 para determinar sus funciones en la migración, apoptosis y proliferación de células endoteliales. La expresión de miR-133a-3p, PSAT1, GSK3β, β-catenina, Ki-67, CyclinD1, Bax y Bcl-2 en las CE se ensayaron mediante RT-qPCR o análisis de transferencia Western. Además, se estableció un modelo de rata IA para detectar los cambios patológicos y la expresión de miR-133a-3p, PSAT1, GSK3β, β-catenina, VEGF y MMP-9 en tejidos IA in vivo. La expresión de miR-133a-3p se relacionó con el número y tamaño de IA. La expresión de MiR-133a-3p había fallecido y la expresión de PSAT1, GSK3β y β-catenina se elevó en IA. El miR-133a-3p restaurado y el PSAT1 reducido aliviaron el cambio patológico; expresión reducida de PSAT1, GSK3β y β-catenina en IA; suprimió la apoptosis y la proliferación y migración avanzadas de las CE de IA, así como la expresión reducida de VEGF y MMP-9 en tejidos de IA in vivo. Nuestro estudio sugiere que la sobreexpresión de miR-133a-3p o la regulación a la baja de PSAT1 restringe el daño de las células endoteliales y avanza la proliferación de células endoteliales mediante la inhibición de la vía GSK3β / β-catenina en IA. MiR-133a-3p podría ser un marcador candidato potencial y una diana terapéutica para IA.
Presentación de la hipótesis
En este estudio, podríamos especular que el eje miR-133a-3p / PSAT1 podría afectar la IA inducida por daño de las células endoteliales a través de la modulación de la vía GSK3β / β-catenina.
Prueba de la hipótesis
Para verificar esta hipótesis, recolectamos muestras clínicas, células endoteliales (CE) del IA humano y establecimos modelos de rata IA para dilucidar las funciones de miR-133a-3p y PSAT1 en el proceso IA.
Implicaciones de la hipótesis
Nuestro estudio confirma nuestra hipótesis de que la sobreexpresión de miR-133a-3p o la regulación a la baja de PSAT1 restringen el daño de las células endoteliales y avanzan la proliferación de células endoteliales mediante la inhibición de la vía GSK3β / β-catenina en IA. Estos hallazgos brindan una nueva perspectiva en una nueva terapia diana para la AI.
Introducción
El aneurisma intracraneal (AI) es un tipo de trastorno cerebrovascular, que se caracteriza por un abultamiento no convencional de la arteria en el cerebro, así como hemorragia subaracnoidea (HSA) causada por ruptura del AI, acompañada de una alta mortalidad y morbilidad [1]. Como es una enfermedad destructiva, la patogenia de la AI no ha sido aclarada [2]. La AI es una forma familiar rara, pero generalmente se piensa que es el resultado de una lesión vascular adquirida causada por hipertensión, tabaquismo y otros factores de riesgo tradicionales [3]. Se han utilizado espirales endovasculares o clips microquirúrgicos para prevenir la futura rotura de un aneurisma no roto en pacientes con alto riesgo de rotura [4]. Aunque se ha logrado un progreso significativo en la operación quirúrgica AI, todavía se presenta una mala recuperación posoperatoria en los pacientes con AI [5]. La difícil situación del tratamiento IA hace necesario seguir explorando el mecanismo y encontrar una nueva estrategia terapéutica.
Los microARN (MiARN) son una clase de ARN no codificantes que modulan la expresión de genes diana mediante la inhibición de la traducción en el nivel postranscripcional o mediando la degradación del ARNm [6]. Se ha revelado que miR-133a-3p como inhibidor tumoral en varias neoplasias malignas y la sobreexpresión de miR-133a-3p pueden inhibir el crecimiento de células de cáncer colorrectal (CCR) [7]. Un estudio anterior ha sostenido que a la luz del bloqueo de la glutaminólisis mediada por autofagia, miR-133a-3p suprime aún más la metástasis y el crecimiento del cáncer gástrico [8]. Además, un estudio ha presentado que miR-133a-3p participa en la regulación del desarrollo cardíaco y la hipertrofia cardíaca [9]. Un estudio ha demostrado que la regulación positiva de miR-195-5p disminuye la angiogénesis y la resistencia al cisplatino en el cáncer de ovario mediante la supresión de la vía de señalización GSK3β / β-catenina dependiente de la fosfoserina aminotransferasa 1 (PSAT1) [10]. Otro estudio ha informado que miR-365 reprime la invasión celular y el crecimiento en el carcinoma de células escamosas de esófago (ESCC) mediante la modulación de PSAT1 [11]. PSAT1 es una enzima implicada en la biosíntesis de serina; originalmente se purifica a partir del cerebro de las ovejas y tiene niveles elevados en muchos tejidos [12]. Se ha documentado que PSAT1 media la progresión del ciclo celular en el cáncer de mama a través de la modulación de la vía de señalización de GSK3β / β-catenina [13]. Liu y col. también señala que PSAT1 ejerce una función sobre el desarrollo de ESCC y pronostica una escasa supervivencia; por lo tanto, puede ser un objetivo prometedor para las terapias contra el cáncer [14]. Dados los análisis antes mencionados, se esperaba que este estudio hiciera contribuciones a un nuevo enfoque para el papel funcional del eje miR-133a-3p / PSAT1 / GSK3β / β-catenina en IA.
Materiales y métodos
Declaración de ética
El estudio fue aprobado por la Junta de Revisión Institucional del Primer Hospital Docente de la Universidad de Medicina Tradicional China de Tianjin. Todos los participantes firmaron un documento de consentimiento informado. Todos los experimentos con animales se compararon con la Guía para el cuidado y uso de animales de laboratorio de los comités internacionales.
Temas de estudio
De enero de 2016 a marzo de 2018, se seleccionaron los casos de HSA por AI tratados en la neurocirugía del Primer Hospital Docente de la Universidad de Medicina Tradicional China de Tianjin. Las muestras patológicas de 75 casos de AI obtenidas por microcirugía fueron recolectadas y clasificadas como grupo AI, incluyendo 29 hombres y 46 mujeres de 31 a 55 años, con una edad promedio de 44,98 ± 6,79 años. Los pacientes con trauma cerebral tratados en neurocirugía al mismo tiempo en el Primer Hospital Docente de la Universidad de Medicina Tradicional China de Tianjin fueron seleccionados como grupo de control. Además, se acumularon 75 casos de tejidos arteriolares intracraneales normales por operación traumática o descompresión interna, incluidos 43 hombres y 32 mujeres de 34 a 56 años, con una edad promedio de 48,14 ± 8,68 años. Los pacientes fueron excluidos si tenían antecedentes de hipertensión, diabetes o tumores. No hubo una diferencia marcada en el género y la edad entre el grupo IA y el grupo de control (tanto P > 0,05).
Tratamiento y conservación de muestras
Después de la resección quirúrgica, algunas de las muestras en dos grupos se fijaron con formaldehído, se deshidrataron con alcohol en gradiente de concentración baja a alta y se embebieron con parafina. Luego, las muestras se cortaron en rodajas para tinción con hematoxilina-eosina (HE) y tinción inmunohistoquímica. Algunas muestras se colocaron rápidamente en tanques de nitrógeno líquido y luego se transfirieron a un refrigerador criogénico a -80 ° C para la detección del análisis de transferencia Western y la reacción en cadena de la polimerasa cuantitativa con transcripción inversa (RT-qPCR). Algunas muestras se fijaron con glutaraldehído para la observación con microscopio electrónico y algunas muestras se utilizaron para el aislamiento de CE.
Observación con microscopio electrónico
Las muestras se fijaron con glutaraldehído al 3% y luego se volvieron a fijar con tetróxido de osmio al 1%. Las muestras se deshidrataron con acetona, se incrustaron con Epon812 y se cortaron en secciones semifinas con un grosor de 3 µm. Por último, las muestras se tiñeron dos veces con acetato de uranilo y citrato de plomo y se observaron con un microscopio electrónico de transmisión H-600IV (Hitachi, Tokio, Japón).
Tinción HE
Las secciones de parafina preparadas se hornearon a 60 ° C durante 30 min. Una vez completados los pasos anteriores, los cortes de tejido se fijaron en xileno, se deshidrataron con alcohol absoluto en gradiente y se limpiaron con solución salina tamponada con fosfato (PBS). Los cortes de tejido se tiñeron con hematoxilina, se trataron con amoniaco durante unos segundos, se tiñeron con eosina durante 2 min, se deshidrataron y aclararon. Luego, las rodajas de tejido se gotearon con goma neutra y se sellaron con un cubreobjetos. Finalmente, se adoptó un microscopio (Nikon, Tokio, Japón) para observar y registrar.
Tinción inmunohistoquímica
El kit de inmunohistoquímica fue producido por Zymed Laboratories (San Francisco, CA, EE. UU.). Las rodajas de parafina se desparafinaron e hidrataron, y las rodajas de parafina se sumergieron en una solución de xileno durante 5 min x 3 veces. Las rodajas se colocaron en alcohol absoluto al 100% durante 3 min x 2 veces y luego se remojaron en alcohol al 95-75% durante 3 min. Después del desparafinado, las rodajas se incubaron con peróxido de hidrógeno al 3% durante 15 min para eliminar la actividad de la peroxidasa endógena. Se gotearon las rodajas con solución de bloqueo y se incubaron con solución de trabajo de suero de cabra normal durante 15 min, luego se sondaron con anticuerpo primario contra metaloproteasa de matriz (MMP) -9 (5 μg / mL) y factor de crecimiento endotelial vascular (VEGF) (1:250 , Abcam, Cambridge, MA, EE. UU.) (PBS para control negativo (NC)) y se incubó durante 1 a 2 h. Los cortes se volvieron a sondar con solución de trabajo de anticuerpo secundario de biotinilación durante 30-60 min. Las rodajas se añadieron con fluido de trabajo de estreptavidina / peroxidasa marcado con peroxidasa de rábano picante, se gotearon con una solución de diaminobencidina (DAB) recién preparada, se contratiñeron y se bloquearon. La imagen fue conocida por el sistema de imágenes Nikon SPOT FlexTM. El área de expresión de la proteína MMP-9 y VEGF se midió mediante un software de análisis cuantitativo inmunohistoquímico. Se detectaron aleatoriamente cinco campos visuales de alta potencia en el área de acumulación de células positivas en cada muestra, y la absorbancia promedio de cada pantalla se utilizó como valor medio para el análisis estadístico.
Aislamiento y cultivo de CE
Las CE se aislaron de tejidos arteriolares intracraneales normales y tejidos IA y se cultivaron. El tejido se cortó en 3 mm 2 fragmentos y se incubaron durante 25 min en colagenasa B al 0,1% / dispasa al 0,1% (Roche, Basilea, Suiza). El tejido se separó, se trituró durante 2 min con una pipeta de 2 ml y se filtró con un colador de 100 µm (BD Biosciences, NJ, EE. UU.) Para aislar las CE. La suspensión celular se centrifugó y luego se resuspendió en un medio de cultivo MV2 que contenía factores de crecimiento y suero bovino fetal al 20% (PromoCell, Heidelberg, Alemania). A continuación, las células se sembraron en placas recubiertas con fibronectina (Sigma Aldrich Inc., St. Louis, MO, EE. UU.) Con una densidad de 10 4 células / cm 2 (1 μg / cm 2 ) y se cultivó durante 1 día con 5% CO 2 . El día después de la siembra, las células se enjuagaron con PBS para eliminar las células no unidas y se colocaron en un medio nuevo. Cuando alcanzaron aproximadamente un 80-100% de confluencia, los cultivos se expusieron a inmunoseparación por perlas recubiertas de aglutinina I (UEA) de Ulex europaeus (Vector Laboratories, Ltd., Peterborough, Reino Unido) (Dynabeads M-450 Tosylactivated, Oxoid, Hampshire, Reino Unido) para obtener EC puros. Las CE unidas a las perlas recubiertas de lectina se acumularon con un concentrador de partículas magnéticas mientras que las células no unidas se eliminaron mediante lavado con medio basal dos veces. Las células positivas para UEA se resuspendieron en medio de cultivo y se sembraron en placas recubiertas de fibronectina para mejorar su adhesión y crecimiento. Los cultivos se volvieron confluentes en 4 a 6 días.
Identificación de CE
Las CE se identificaron mediante tinción inmunocitoquímica con un anticuerpo CD31 de superficie celular y un antígeno relacionado con el factor FVII. Las células se limpiaron dos veces con PBS, se fijaron con paraformaldehído al 4%, se incubaron con H 2 al 3%. O 2 durante 10 a 15 min para eliminar la actividad de peroxidasa endógena y luego se incuban con Triton X-100 al 0,1% durante 10 min hasta las células perforadas. Se gotearon las células con anticuerpo primario específico:Factor VII (1:200), CD31 (1:400, Roche, Basilea, Suiza) y se incubaron a 4 ° C durante la noche. Luego, las células se gotearon con inmunoglobulina G (1:50) marcada con anticuerpo secundario de peroxidasa de rábano picante. Las células se incubaron a 37 ° C durante 45 min y se revelaron mediante DAB evitando la luz durante 4 min. Luego, se terminó el desarrollo del color con el agua destilada y se observó la fotografía al microscopio. Las células se observaron bajo el microscopio de diferencia de fase invertida de fluorescencia, y las células positivas y el número total de células se contaron al azar a partir de 10 campos visuales. La tasa de células de tinción positiva =(el número de células positivas / el número total de células) × 100%. Se estableció el grupo NC correspondiente y el anticuerpo primario se reemplazó por PBS, y los otros pasos se realizaron como se indicó anteriormente.
Agrupación y transfección de células
Para estudiar los efectos entre miR-133a-3p y PSAT1 en las CE de IA, las CE se agruparon en grupo de control (CE vasculares normales sin ninguna transfección), grupo IA (CE vasculares IA sin ninguna transfección), grupo NC imitado (transfectado con miR -133a-3p mimic NC), grupo miR-133a-3p mimic (transfectado con miR-133a-3p mimic), pequeño grupo de ARN interferente (si) -NC (transfectado con si-PSAT1 NC), grupo si-PSAT1 (transfectado con si-PSAT1) y miR-133a-3p mimic + sobreexpresión (oe) -PSAT1 grupo (transfectado con miR-133a-3p mimic y oe-PSAT1). Entre ellos, mimic NC, miR-133a-3p mimic, si-PSAT1, si-NC y oe-PSAT1 fueron ideados y compuestos por GenePharma Co., Ltd. (Shanghai, China). La transfección se realizó estrictamente de acuerdo con las instrucciones de Lipofectamine TM 2000 reactivo de transfección (Thermo Fisher Scientific, MA, EE. UU.).
Citometría de flujo
El medio de la placa de cultivo se desechó y las células se aclararon dos veces con PBS. Las células se separaron con tripsina al 0,25%, se centrifugaron a 800 rpm durante 5 min y se suspendieron con tampón de unión 1 ×, y la densidad celular se ajustó a 1 × 10 7 células / mL. La suspensión celular (100 μL) se incubó con 5 μL de yoduro de propidio (PI, 20 μg / mL) y anexina V-FITC durante 20 min, luego se mezcló con 400 μL de tampón de unión 1x. Se utilizó un citómetro de flujo (clasificador de células BD FACSArial I) para detectar la apoptosis celular en 1 h. Los resultados fueron que el cuadrante inferior izquierdo (Q4) en el mapa de dispersión mostraba células vivas sanas (FITC - / PI - ), el cuadrante inferior derecho (Q3) como las células apoptóticas tempranas (FITC + / PI - ), y el cuadrante superior derecho (Q2) fueron las células apoptóticas y apoptóticas tardías (FITC + / PI + ); tasa de apoptosis =porcentaje de apoptosis temprana (Q3) + porcentaje de apoptosis tardía (Q2).
Ensayo de bromuro de 3- (4, 5-dimetiltiazol-2-Yl) -2, 5-difeniltetrazolio (MTT)
Las células se separaron con tripsina para preparar la suspensión celular. Las células se contaron con un microscopio invertido. La concentración celular se ajustó a 5 × 10 4 células / mL. Las células se sembraron en una placa de cultivo de 96 pocillos. Después de 48 h, las células se incubaron con 20 μL de solución de MTT durante 4 h. Se disolvió MTT en cada pocillo con 150 µl de dimetilsulfóxido. El valor de densidad óptica (DO) de las CE se calculó a la longitud de onda de 570 nm. La tasa de proliferación de CE se calculó a la luz del valor de DO.
Prueba de Scratch
Las células de cada grupo se sembraron en una placa de 24 pocillos con 2 × 10 5 células / pocillo. Se colocaron tres pozos paralelos en cada grupo. Cuando se alcanzó una confluencia de aproximadamente el 90%, se raspó el plano de crecimiento celular con una punta de micropipeta desechable esterilizada de 1 ml; cada pocillo se raspó una vez y la longitud y la profundidad de raspado de cada pocillo fueron consistentes. Después de raspar, se retiraron las células flotantes, se reemplazó el medio de cultivo con uno nuevo y se observó el espaciamiento de raspado bajo el microscopio después de 24 h de cultivo. El área de cicatrización de la herida por arañazo fue enumerada por el software National Instrument Vision Assistant 8.6. La migración celular =área de cicatrización de la herida / área inicial de la herida por raspado × 100%.
Animales de experimentación y establecimiento de modelos de ratas IA
Se seleccionaron ochenta y cuatro ratas Sprague-Dawley (SD) de 7 semanas de edad y con un peso de 180 a 200 g (Centro de Animales de Laboratorio, Academia de Ciencias Médicas Militares, Beijing, China). Las ratas se alojaron en el centro de experimentación animal. Las condiciones de alimentación se controlaron a 22-25 ° C y 50-60% de humedad con luz natural. Todas las ratas se alimentaron en jaulas para ratas estándar con 4 ratas por jaula. Las ratas fueron alimentadas con agua potable urbana y forraje común para ratas. Los cojines se cambiaron cada 3 días y la jaula se lavó y esterilizó. Las ratas IA se modelaron de acuerdo con la referencia [15]. Se identificó un aneurisma roto cuando se presentaron los siguientes síntomas en ratas [16]:1, disminución de la actividad de comer o beber realizada por una pérdida de peso (aproximadamente un 10% de pérdida de peso) durante 24 h; 2, flexión del torso y extremidades anteriores al levantar; 3, caminar de un lado en una postura normal; 4, inclinado hacia un lado en reposo, sin actividad espontánea. Las ratas con estos síntomas fueron sacrificadas 3 meses después de la operación. Se obtuvieron tejidos IA durante la operación y se perfundieron con PBS, y el colorante azul que contenía ácido glutámico se perfundió en la arteria cerebral.
Tratamiento e intervención de ratas IA
Las 84 ratas anteriores se separaron aleatoriamente en 7 grupos con 12 ratas en cada grupo. Los métodos de tratamiento fueron los siguientes:grupo normal (no se realizó ningún modelo); Grupo IA (estereotáctico inyectado con 100 μl de mezcla de PBS y Lipofectamine 2000); imitar el grupo NC (inyección estereotáctica con 100 µl de mezcla de miR-133a-3p imitar NC y Lipofectamine 2000); grupo imitador de miR-133a-3p (inyección estereotáctica con 100 µl de mezcla de imitador de miR-133a-3p y Lipofectamine 2000); grupo si-NC (inyección estereotáctica con 100 μL de mezcla de si-PSAT1 NC y Lipofectamine 2000); grupo si-PSAT1 (inyección estereotáctica con 100 μL de mezcla de si-PSAT1 y Lipofectamine 2000); y miR-133a-3p mimic + oe-PSAT1 grupo (inyección estereotáctica con 100 µl de mezcla de miR-133a-3p mimic y oe-PSAT1 y Lipofectamine 2000). Todas las inyecciones anteriores se realizaron una vez al día, y estas ratas se criaron en un laboratorio de animales libres de patógenos específicos (SPF) durante 12 semanas. Después de 12 semanas, las ratas de cada grupo se anestesiaron y se abrió la cavidad torácica como se describió anteriormente. Desde el ventrículo izquierdo que se intuba hacia la aorta, se liberó sangre cortando la cava. Al mismo tiempo, se perfundieron 30 ml de solución salina que contenía heparina sódica (37ºC) a través del conducto, y luego, se inyectó lentamente poliformaldehído al 10% / tampón fosfato 0,1 M (pH 7,4) en el cerebro a través del conducto. Una vez fijada la perfusión, se abrió el cerebro. El anillo arterial en la base del cráneo se separó y se extrajo bajo el microscopio quirúrgico, se observaron los cambios de los aneurismas bajo el microscopio y se relevaron las características patológicas. El imitador NC, miR-133a-3p imitador, si-NC, si-PSAT1 y oe-PSAT1 fueron compuestos por Shanghai Sangon Biotechnology Co., Ltd. (Shanghai, China).
Detección de hemodinámica
El caudal de sangre al final de la arteria carótida común izquierda de ratas se evaluó antes de los 3 días de operación y 12 semanas después del tratamiento de intervención. El método fue el siguiente:las ratas se colocaron en el marco animal de la máquina de anestesia para la inhalación y se ajustaron los parámetros de flujo. Después de que las ratas respiraran de manera estable y no hubo una reacción obvia cuando se tocó la cola de las ratas, las ratas se fijaron en la mesa de operaciones experimental con una goma elástica. El pelo del cuello de la rata se afeitó con una afeitadora eléctrica. Se encendió el detector de ultrasonido Doppler color, se midió la velocidad del flujo sanguíneo al final de la arteria carótida común izquierda y se registraron los datos después de que la sonda se untó con el agente de acoplamiento apropiado. Después de la medición, las ratas se volvieron a colocar con cuidado en la jaula para mantener el tracto respiratorio despejado hasta que las ratas se despertaran después de la anestesia.
RT-qPCR
El ARN total se extrajo sobre la base de un kit de extracción de ARN total simple de ARN (TIANGEN Biotechnology Co., Ltd., Beijing, China). El ARN de alta calidad se confirmó mediante análisis ultravioleta y electroforesis de desnaturalización con formaldehído, y el ARN se transcribió a la inversa en ADN complementario mediante el kit de reactivos PrimeScript RT. La reacción de PCR se llevó a cabo con SYBR Permix Ex Taq II (Takara, Dalian, Liaoning, China). Los cebadores de PCR fueron ideados y combinados por Beijing ComWin Biotech Co., Ltd. (Beijing, China) (Tabla 1). Se seleccionó U6 como parámetro interno para miR-133a-3p, PSAT1, GSK3β, β-catenina, Bax, Bcl-2, Ki-67 y CyclinD1 con gliceraldehído fosfato deshidrogenasa (GAPDH) como parámetro interno. Los datos se midieron con 2 −ΔΔCt .
Análisis de Western Blot
Las proteínas totales se extrajeron de células y tejidos, y las muestras de proteínas se cuantificaron mediante el kit de ensayo de proteínas de ácido bicinconínico (Instituto de Biotecnología Beyotime, Shanghai, China). Las muestras se mezclaron con 1/4 de volumen de tampón de muestra 5X y se hirvieron durante 5 min. El gel de separación al 10% y el gel concentrado al 5% se seleccionaron para electroforesis. La membrana se incubó en leche desnatada en polvo al 5% durante 60 min. La membrana se añadió con el anticuerpo primario PSAT1 (1:500), GSK3β (1:500), β-catenina (1:5000), Bax (1:1000), Bcl-2 (1:1000), CyclinD1 (1:200), Ki-67 (1:5000), MMP-9 (1 μg / ml), VEGF (1:1000) (todos de Abcam, Cambridge, MA, EE. UU.). Luego, se incubó la membrana con anticuerpo secundario (1:2000) durante 60 min. La membrana se sumergió en la solución de reacción de electroquimioluminiscencia (Beyotime Institute of Biotechnology, Shanghai, China) durante 1 min, luego se cubrió con una envoltura de alimentos después de retirar el fluido. La membrana se expuso mediante rayos X y se observó el resultado después del revelado y la fijación. Se utilizó GAPDH (1:10000, Abcam) como control de carga y la imagen de la proteína se analizó con el software ImageJ2x.
Ensayo génico indicador de luciferasa dual
La relación objetivo entre miR-133a-3p y PSAT1 y el sitio de unión entre miR-133a-3p y la región no traducida 3 'de PSAT1 (3'UTR) fueron pronosticadas por un sitio web de bioinformática (https://cm.jefferson.edu/rna22). / Precalculado /). La secuencia de la región promotora 3'UTR de PSAT1 que contiene el sitio de unión de miR-133a-3p se amplificó y se clonó en el plásmido de luciferasa básica pGL3 (Takara Bio Inc., Otsu, Shiga, Japón) para construir el plásmido de tipo salvaje (WT) ( PSAT1-WT) de PSAT1 3′UTR, mientras que el plásmido recombinante PSAT1-MUT mutante (MUT) se formuló mutando el sitio de unión miR-133a-3p en PSAT1-WT con un kit de mutación puntual (Takara Bio Inc., Otsu, Shiga , Japón). Las CE vasculares en la fase de crecimiento logarítmico se sembraron en una placa de 96 pocillos. Cuando la confluencia alcanza aproximadamente el 70%, los plásmidos PSAT1-WT y PSAT1-MUT se mezclaron con plásmidos imitadores NC y miR-133a-3p mediante Lipofectamine 2000 y se cotransfectaron en EC vasculares. Las células se recogieron y lisaron 48 h después de la transfección, y la actividad de luciferasa se verificó mediante un kit de detección de luciferasa (Promega Corporation, Madison, WI, EE. UU.).
Análisis estadístico
Todos los datos fueron explicados por el software SPSS 21.0 (IBM Corp., Armonk, NY, EE. UU.). Los datos de enumeración se indicaron mediante tasa o porcentaje, y el análisis se determinó mediante la prueba de chi-cuadrado o la prueba de Fisher. Los datos de medición sometidos a distribución normal se transmitieron mediante media ± desviación estándar. t realizó la comparación entre dos grupos prueba, mientras que la comparación entre múltiples grupos se analizó mediante análisis de varianza unidireccional (ANOVA) seguido de la prueba post hoc de Tukey. A P valor <0,05 se consideró significativo.
Resultados
Datos generales de pacientes con AI
Como se muestra en la Tabla 2, se compararon los datos generales del grupo IA y el grupo de control. La información específica se enumera en la Tabla 2.
La expresión de miR-133a-3p está relacionada con el número y tamaño de IA
Al analizar la relación entre la expresión de miR-133a-3p y las características clínico-patológicas de IA, se detalló en la Tabla 3 que a la luz de la expresión relativa promedio de miR-133a-3p en IA, 75 casos de IA se distribuyeron en dos grupos:grupo de alta expresión miR-133a-3p ( n =47) y miR-133a-3p grupo de baja expresión ( n =28). La relación entre miR-133a-3p y diferentes parámetros clínico-patológicos se analizó estadísticamente mediante la prueba de chi-cuadrado o la prueba de Fisher. Los resultados mostraron que la expresión de miR-133a-3p no estaba relacionada con la edad, el sexo, la forma y la posición del aneurisma (todos P > 0.05), pero asociado con el número y tamaño del aneurisma (tanto P <0.05).
La expresión de MiR-133a-3p está disminuida y la expresión de PSAT1, GSK3β y β-catenina aumenta en los tejidos IA
La expresión de miR-133a-3p en IA se determinó mediante RT-qPCR, y los resultados revelaron que en relación con los tejidos arteriolas intracraneales normales (el grupo de control), la expresión de miR-133a-3p se redujo en los tejidos IA (el Grupo IA) ( P <0.05) (Figura 1a). El análisis de RT-qPCR y Western blot manifestó que la expresión de PSAT1, GSK3β y β-catenina aumentó en los tejidos IA en relación con la de los tejidos de arteriolas intracraneales normales (todos P <0.05) (Fig. 1a – c).
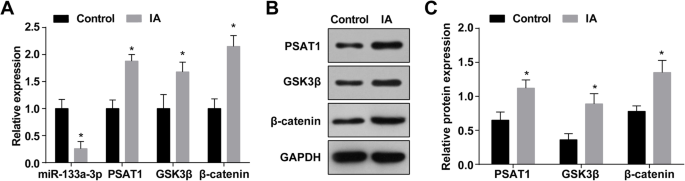
La expresión de MiR-133a-3p disminuye y la expresión de PSAT1, GSK3β y β-catenina aumenta en tejidos IA. un Expresión de miR-133a-3p, PSAT1, GSK3β y β-catenina en el grupo IA y el grupo normal. b Banda de proteína de expresión de PSAT1, GSK3β y β-catenina. c Expresión de proteínas de PSAT1, GSK3β y β-catenina en el grupo IA y el grupo normal mediante ensayo de transferencia Western. n =75, * P <0,05 frente al grupo de control. Los datos de medición se representaron como media ± desviación estándar, y las comparaciones entre dos grupos se realizaron mediante una muestra independiente t prueba
Cambios patológicos de aneurisma y expresión de MMP-9 y VEGF en tejidos IA
Al observar directamente el tejido de las arteriolas intracraneales normales y los tejidos IA, se presentó que en el grupo de control, los vasos en los tejidos arteriales eran de color rojo brillante y no se encontraron placas ateroscleróticas obvias ni trombos laterales en la luz. El tumor de los tejidos del aneurisma del grupo IA era mayoritariamente marrón o rojo oscuro, y el aspecto era escrotiforme o fusiforme, y la textura era mayoritariamente dura. Cuando se abrió el tumor, aparecieron placas ateroscleróticas de color blanco o rojo oscuro en la pared del tumor de algunas muestras de tumor, que eran de forma plana, redonda u ovalada. En algunas muestras de tumor, había un trombo mural en la cavidad tumoral y la textura del trombo era blanda. El grosor de la pared del tumor se adelgazó gradualmente desde el cuello del tumor, algunos de los cuales solo tenían una membrana de fibra delgada en la parte superior del tumor, y algunos de ellos incluso se habían roto. La grieta del aneurisma roto estaba ubicada en o cerca de la parte superior del tumor.
La tinción HE mostró que bajo el microscopio óptico, el grosor de la pared de los tejidos de las arteriolas intracraneales normales era uniforme; la estructura anatómica de las capas interna, media y externa estaba clara e intacta; la morfología de las células en cada capa era normal; el sarcolema de las células adyacentes a menudo se formó de forma estrecha; y las células inflamatorias de la pared eran raras. En el grupo IA, las protuberancias locales formadas en la cavidad vascular lateral distal en la parte superior de la bifurcación de la arteria intracraneal en la pared del aneurisma se volvieron obtusas y más pequeñas, y se perdieron las CE locales. Un pequeño número de muestras mostró una migración de la capa de células de músculo liso a la capa de la íntima y una proliferación de células de la íntima miogénica. Las CE disminuyeron o incluso desaparecieron. La capa de células endoteliales estaba compuesta por células miointimales hiperplásicas y CE dispuestas linealmente, o por CE apoptóticas y células sanguíneas adheridas a la luz. Su vacuola degeneró y se presentó con interrupción de continuidad. Algunos de ellos se despegaron junto con la membrana basal y se aumentaron las fibras de colágeno de la íntima. La aterosclerosis cambió y la pared de la arteriola era obviamente más delgada y estaba llena de una gran cantidad de tejidos conectivos en escalera. Se observó infiltración celular inflamatoria y difusión parcial en todas las capas, principalmente en las membranas media y externa. Se observaron depósitos de lípidos y cristales de colesterol en algunas células. Algunas de las paredes del tumor se adelgazaron completa o localmente y se expandieron hacia afuera (Fig. 2a, b).
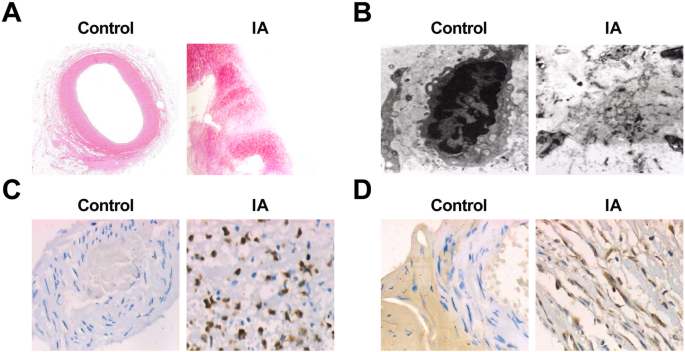
Cambios patológicos del aneurisma y expresión de MMP-9 y VEGF en IA. un Cortes de tejido de arteriolas intracraneales normales en el grupo de control con tinción HE (× 10). b Secciones de tejido IA teñidas con HE (× 10). c Ultraestructura de los tejidos de las arteriolas intracraneales normales en el grupo de control bajo un microscopio electrónico (× 10,000). d Ultraestructura de los tejidos IA bajo un microscopio electrónico (× 10,000). e Expresión de MMP-9 en el grupo control y el grupo IA mediante tinción inmunohistoquímica (× 200). f Expresión de VEGF en el grupo de control y el grupo IA mediante tinción inmunohistoquímica (× 200)
Las secciones de los tejidos de las arteriolas intracraneales normales y los tejidos IA se observaron mediante un microscopio electrónico, y se realizó que en los tejidos de las arteriolas intracraneales normales, las fibras de la matriz de la pared vascular cerebral se pudieran ver claramente y no hubo lesión endotelial, picnosis celular. , o degeneración. En los tejidos IA, se observó lesión obvia de las células endoteliales, picnosis celular o degeneración de las vacuolas, disminuyó el número de células del músculo liso medio, apareció la mayor parte de la picnosis del núcleo y se pudo observar agregación de cromatina y cuerpos apoptóticos. Algunas células mostraron hinchazón de las mitocondrias y desaparición de la estructura interna normal. La matriz extracelular que formaba el citoesqueleto estaba borrosa y mostraba flóculos amorfos. Había muchos fragmentos en las partes faltantes de las células (Fig. 2c, d).
Se utilizó tinción inmunohistoquímica para probar la expresión de MMP-9 y VEGF, y los resultados revelaron que no había expresión de MMP-9 y VEGF en 75 casos del grupo de control. Existieron 60 casos de expresión positiva de MMP-9 en los 75 casos de muestras IA. La expresión positiva de MMP-9 apareció en las membranas interna y externa de la pared IA, pero la expresión no fue uniforme. La expresión positiva se caracterizó principalmente por un citoplasma amarillo pardusco. La expresión positiva de VEGF fue de 66 casos en 75 casos de muestras IA. En la pared de IA, hubo una alta expresión positiva en las membranas media y externa y una baja expresión positiva en la íntima. La expresión positiva también se caracterizó principalmente por un citoplasma amarillo pardusco (Fig. 2e, f). La expresión de MMP-9 y VEGF en los dos grupos se muestra en la Tabla 4.
Identificación de CE vasculares
La expresión de factor VIII y CD31 en CE se analizó mediante tinción inmunohistoquímica. Los resultados informaron que las CE vasculares reaccionaron positivamente a los anticuerpos antigénicos relacionados con el factor VIII y CD31, y la tasa positiva fue del 95%. Además, había una gran cantidad de partículas marrones en el citoplasma, y el quinto pasaje de células con tinción marrón fue dramáticamente más alto que el pasaje primario de células (Fig. 3a, b).
Las CE vasculares reaccionan positivamente a los anticuerpos antigénicos relacionados con FVIII y CD31. un Identificación de CE por CD31. b Identificación de CE por FVII
La regulación al alza de miR-133a-3p y la regulación a la baja de PSAT1 suprimen la apoptosis y la proliferación y migración avanzadas de CE en IA
Se adoptaron citometría de flujo, RT-qPCR y análisis de inmunotransferencia de tipo Western para observar la apoptosis y la expresión de Bax y Bcl-2 en las CE de IA después de tratarse con miR-133a-3p mimic o si-PSAT1. Se indicó que en comparación con el grupo de control, la tasa de apoptosis de las células y la expresión de Bax se elevó en el grupo IA y la expresión de Bcl-2 disminuyó (todas P <0,05). La apoptosis celular y la expresión de Bax y Bcl-2 en el grupo IA, el grupo NC mímico y el grupo si-NC no tuvieron cambios significativos (todos P > 0,05). En comparación con el grupo si-NC y el grupo NC imitador, se suprimió la tasa de apoptosis de las células en el grupo si-PSAT1 y el grupo imitador miR-133a-3p, se redujo la expresión de Bax y se elevó la expresión de Bcl-2 ( todos P <0,05). Frente al grupo imitador de miR-133a-3p, la tasa de apoptosis y la expresión de Bax mejoraron, y la expresión de Bcl-2 se redujo en el grupo imitador de miR-133a-3p + oe-PSAT1 (todos P <0.05) (Fig. 4a – e).
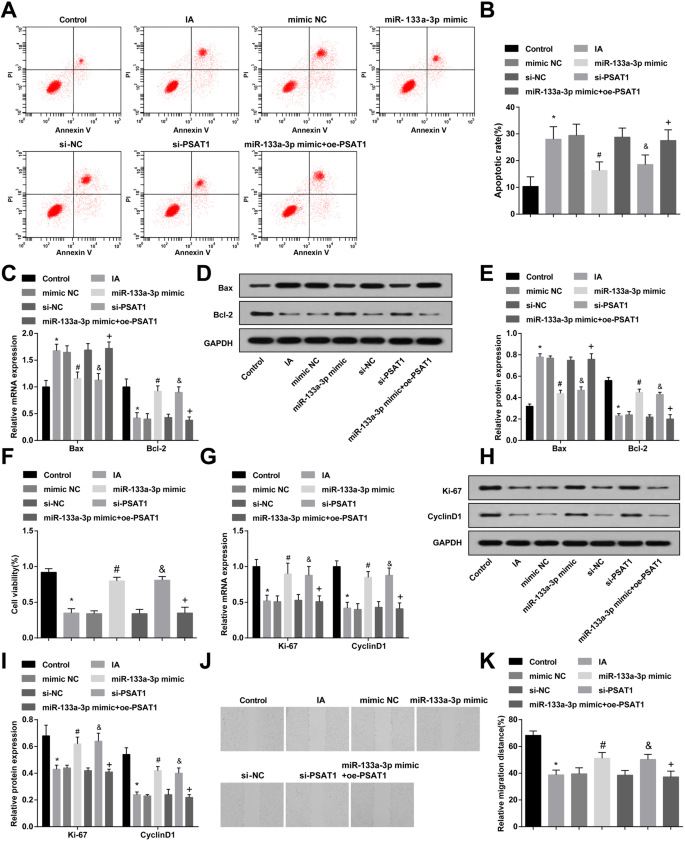
El miR-133a-3p altamente expresado y el PSAT1 de baja expresión inhiben la apoptosis y promueven la proliferación y migración de las CE de IA. un Detección de apoptosis de CE mediante citometría de flujo. b Detección de la tasa de apoptosis de las CE en cada grupo. c Expresión de Bax y Bcl-2 en CE detectadas por RT-qPCR. d Banda proteica de expresión de Bax y Bcl-2. e Expresión de la proteína Bax y Bcl-2 en las CE detectadas mediante análisis de transferencia Western. f Se utilizó el ensayo MTT para detectar la actividad de proliferación de las CE en cada grupo. g Se usó RT-qPCR para detectar la expresión de Ki-67 y CyclinD1 en cada grupo de EC. h Banda de proteína de expresión de Ki-67 y CyclinD1. yo Expresión de la proteína Ki-67 y CyclinD1 en las CE detectadas mediante análisis de transferencia Western. j Detección de la migración de CE en cada grupo mediante prueba de rayado. k Resultados estadísticos de la migración de células endoteliales en cada grupo. N =3, * P <0,05 frente al grupo de control. # P <0,05 frente al grupo NC mímico. & P <0,05 frente al grupo si-NC. + P <0,05 frente al grupo mímico de miR-133a-3p. Los datos de medición se representaron como media ± desviación estándar; Los datos se evaluaron mediante un análisis de varianza unidireccional seguido de la prueba post hoc de Tukey
Se utilizaron el ensayo MTT, RT-qPCR y análisis de transferencia Western para observar la proliferación y la expresión de Ki-67 y CyclinD1 en las CE de IA después de tratarlas con miR-133a-3p mimic o si-PSAT1. Se demostró que, en contraste con el grupo de control, la actividad de proliferación y la expresión de Ki-67 y CyclinD1 se redujeron en el grupo IA (todos P <0,05). No hubo diferencias significativas en la actividad de proliferación y la expresión de Ki-67 y CyclinD1 del grupo IA, el grupo NC imitado y el grupo si-NC (todos P > 0,05). En relación con el grupo si-NC y el grupo imitador NC, la actividad de proliferación y la expresión de Ki-67 y CyclinD1 aumentaron en el grupo si-PSAT1 y el grupo mimético miR-133a-3p (todos P <0,05). En comparación con el grupo mimético de miR-133a-3p, la actividad de proliferación y la expresión de Ki-67 y CyclinD1 se redujeron en el grupo mimético de miR-133a-3p + oe-PSAT1 (todos P <0.05) (Fig. 4f, i).
La migración de las CE en cada grupo después del tratamiento con miR-133a-3p mimic o si-PSAT1 durante 24 h se observó mediante la prueba de raspado. Se reveló que la migración de células en el grupo IA se inhibió en relación con la del grupo de control ( P <0,05). No hubo un cambio notable en la migración celular del grupo IA, el grupo si-NC y el grupo NC mímico (todos P > 0,05). En comparación con el grupo si-NC y el grupo NC imitador, la migración celular en el grupo si-PSAT1 y el grupo imitador miR-133a-3p fue elevada (ambos P <0,05). Frente al grupo mimético de miR-133a-3p, la migración celular se redujo en el grupo mimético de miR-133a-3p + oe-PSAT1 ( P <0.05) (Fig. 4j, k).
MiR-133a-3p restaurado y PSAT1 empobrecido reducen la expresión de PSAT1, GSK3β y β-catenina en las CE de IA
Se usó RT-qPCR para detectar la expresión de miR-133a-3p en las CE de IA; Se obtuvo que, en comparación con el grupo de control, la expresión de miR-133a-3p en el grupo IA se redujo ( P <0,05). La expresión de miR-133a-3p en el grupo IA, el grupo imitador NC y el grupo si-NC no cambió notablemente ( P > 0,05). La expresión de MiR-133a-3p en el grupo imitador de miR-133a-3p se incrementó en relación con la del grupo imitador NC ( P <0,05). En contraste con el grupo si-NC, no hubo cambios claros en la expresión de miR-133a-3p en el grupo si-PSAT1 ( P > 0,05). Frente al grupo imitador de miR-133a-3p, la expresión de miR-133a-3p no mostró diferencias significativas en el grupo imitador de miR-133a-3p + oe-PSAT1 ( P > 0.05) (Fig. 5a).
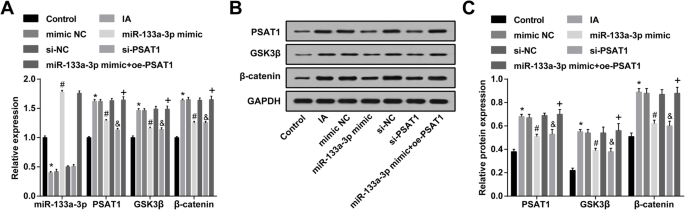
La regulación al alza de miR-133a-3p y la regulación a la baja de PSAT1 disminuyen la expresión de PSAT1, GSK3β y β-catenina en las CE de IA. un Expresión de miR-133a-3p, PSAT1, GSK3β y β-catenina en las CE detectadas por RT-qPCR. b Bandas de proteínas de PSAT1, GSK3β y β-catenina. c Expresión de la proteína PSAT1, GSK3β y β-catenina en las CE en cada grupo detectado por análisis de transferencia Western. N =3, * P <0,05 frente al grupo de control. # P <0,05 frente al grupo NC mímico. & P <0,05 frente al grupo si-NC. + P <0,05 frente al grupo mímico de miR-133a-3p. Los datos de medición se representaron como media ± desviación estándar, y los datos se evaluaron mediante un análisis de varianza de una vía seguido de la prueba post hoc de Tukey
La expresión de PSAT1, GSK3β y β-catenina en las CE de IA se ensayó mediante análisis de transferencia Western y RT-qPCR. Se indicó que en relación con el grupo control, la expresión de PSAT1, GSK3β y β-catenina en el grupo IA se elevó (todos P <0,05). La expresión de PSAT1, GSK3β y β-catenina en el grupo IA, el grupo imitador NC y el grupo si-NC no cambió drásticamente (todos P > 0,05). La expresión de PSAT1, GSK3β y β-catenina en el grupo imitador de miR-133a-3p y el grupo si-PSAT1 se degradó en relación con la del grupo imitador NC y el grupo si-NC (todos P <0,05). En relación con el grupo de imitadores de miR-133a-3p, la expresión de PSAT1, GSK3β y β-catenina fue elevada en el grupo de imitadores de miR-133a-3p + oe-PSAT1 (todos P <0.05) (Fig. 5a – c).
La regulación al alza de miR-133a-3p y la regulación a la baja de PSAT1 alivian los cambios patológicos de los tejidos IA
Al probar los cambios hemodinámicos de las ratas después del modelado, monitoreamos la velocidad del flujo sanguíneo de las ratas en cada grupo 3 días antes de la operación y 12 semanas después del tratamiento de intervención. Se realizó que no había una diferencia obvia en la velocidad del flujo sanguíneo en cada grupo 3 días antes de la operación ( P > 0,05). Después de 12 semanas de intervención, la velocidad del flujo sanguíneo de las ratas del grupo IA disminuyó en relación con la del grupo normal ( P <0,05). No hubo una diferencia clara en el grado de disminución de la velocidad del flujo sanguíneo en el grupo IA, el grupo imitador NC, el grupo si-NC y el grupo miR-133a-3p imitador + oe-PSAT1 (todos P > 0,05). En comparación con el grupo si-NC y el grupo NC imitador, la velocidad del flujo sanguíneo se incrementó en el grupo imitador miR-133a-3p y el grupo si-PSAT1 (ambos P <0,05). En contraste con el grupo mimético de miR-133a-3p, la velocidad del flujo sanguíneo disminuyó en el grupo mimético de miR-133a-3p + oe-PSAT1 ( P <0.05) (Fig. 6a).
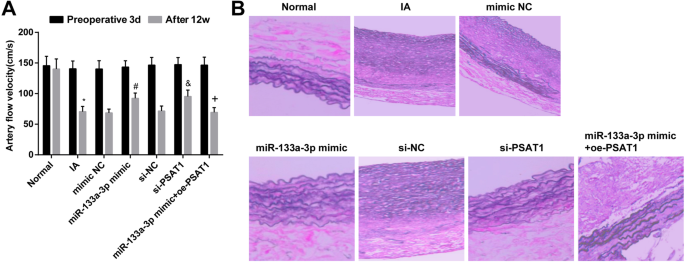
El miR-133a-3p regulado al alza y el PSAT1 regulado a la baja alivian los cambios patológicos de los tejidos IA. un Cambios hemodinámicos en cada momento después de un modelado exitoso en ratas. b Cambios de tejido IA después de la transfección. n =12, * P <0,05 frente al grupo normal. # P <0,05 frente al grupo NC mímico. & P <0,05 frente al grupo si-NC. + P <0,05 frente al grupo mímico de miR-133a-3p. Los datos de medición se representaron como media ± desviación estándar, y los datos se evaluaron mediante un análisis de varianza de una vía seguido de la prueba post hoc de Tukey.
Los cambios de los tejidos IA se verificaron mediante tinción HE. Los resultados mostraron que en el grupo normal, las fibras elásticas en la capa media del tejido vascular intracraneal estaban limpias, apareció la estructura de onda de proteína elástica normal y no hubo rotura ni degradación. En relación con el grupo normal, la luz del tejido vascular intracraneal se agrandó, la estructura en forma de onda de proteína elástica normal desapareció, la fibra elástica en la capa media del vaso de proteína elástica local se rompió y algunas de las fibras elásticas estaban completamente degradado en el grupo IA. No hubo cambios claros en la morfología de los tejidos IA en el grupo si-NC, el grupo imitador NC, el grupo IA y el grupo miR-133a-3p imitador + oe-PSAT1. En contraste con el grupo si-NC y el grupo NC imitador, existía la estructura de onda de la proteína elástica en el tejido vascular intracraneal de ratas en el grupo imitador miR-133a-3p y el grupo si-PSAT1, y la proteína elástica local vascular la estructura estaba levemente desordenada, pero no había fractura ni disolución (Fig. 6b).
MiR-133a-3p altamente expresado y PSAT1 poco expresado reducen la expresión de PSAT1, GSK3β, β-catenina, VEGF y MMP-9 en tejidos IA en vivo
La expresión de miR-133a-3p en tejidos IA in vivo fue probada por RT-qPCR, se sugirió que en relación con el grupo normal, la expresión de miR-133a-3p se redujo en el grupo IA ( P <0,05). La expresión de miR-133a-3p en el grupo IA, el grupo imitador NC y el grupo si-NC no cambió obviamente (todos P > 0,05). La expresión de MiR-133a-3p en el grupo imitador de miR-133a-3p fue elevada en relación con la del grupo imitador NC ( P <0,05). En comparación con el grupo si-NC, no hubo cambios marcados en la expresión de miR-133a-3p en el grupo si-PSAT1 ( P > 0,05). Frente al grupo imitador de miR-133a-3p, la expresión de miR-133a-3p no mostró diferencias significativas en el grupo imitador de miR-133a-3p + oe-PSAT1 ( P > 0.05) (Fig. 7a).
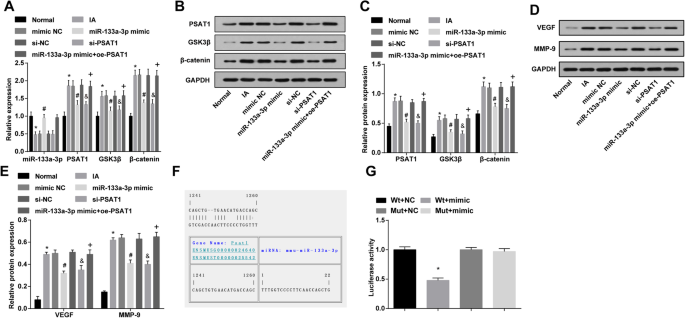
La sobreexpresión de miR-133a-3p y la baja expresión de PSAT1 disminuyen la expresión de PSAT1, GSK3β y β-catenina, VEGF y MMP-9 en tejidos IA in vivo y PSAT1 es un gen diana de miR-133a-3p. un Detección de la expresión de miR-133a-3p, PSAT1, GSK3β y β-catenina en tejidos IA de ratas de cada grupo mediante RT-qPCR. b Bandas de proteínas de PSAT1, GSK3β y β-catenina. c Detección de la expresión de proteínas PSAT1, GSK3β y β-catenina en tejidos IA de ratas en cada grupo mediante análisis de transferencia Western. d Bandas de proteínas de VEGF y MMP-9. e Detección de la expresión de la proteína VEGF y MMP-9 en tejidos IA de ratas en cada grupo mediante análisis de transferencia Western. f Predicción del sitio objetivo de la unión de PSAT1 al miR-133a-3p correspondiente mediante Target Scan. g Resultado del ensayo del gen indicador de luciferasa dual. un - e , n =12; f - g , N =3, * P <0,05 frente al grupo normal / grupo Wt + NC. # P <0,05 frente al grupo NC mímico. & P <0,05 frente al grupo si-NC. + P <0,05 frente al grupo mímico de miR-133a-3p. Los datos de medición se representaron como media ± desviación estándar, y los datos se evaluaron mediante un análisis de varianza de una vía seguido de la prueba post hoc de Tukey
La expresión de PSAT1, GSK3β y β-catenina en tejidos IA in vivo se ensayó mediante análisis de transferencia Western y RT-qPCR. Aparentemente, en contraste con el grupo normal, la expresión de PSAT1, GSK3β y β-catenina en el grupo IA aumentó (todos P <0,05). La expresión de PSAT1, GSK3β y β-catenina en el grupo IA, el grupo imitador NC y el grupo si-NC no cambió notablemente (todos P > 0,05). La expresión de PSAT1, GSK3β y β-catenina en el grupo mímico de miR-133a-3p y el grupo si-PSAT1 disminuyó en comparación con la del grupo NC mímico y el grupo si-NC (todos P <0,05). Frente al grupo imitador de miR-133a-3p, la expresión de PSAT1, GSK3β y β-catenina se incrementó en el grupo imitador de miR-133a-3p + oe-PSAT1 (todos P <0.05) (Fig. 7a – c).
Se utilizó análisis de transferencia Western para verificar la expresión de VEGF y MMP-9 en tejidos IA in vivo; Los resultados percibieron que, en comparación con el grupo normal, la expresión de VEGF y MMP-9 en el grupo IA mejoró (tanto P <0,05). La expresión de VEGF y MMP-9 en el grupo IA, imita el grupo NC y el grupo si-NC no cambió obviamente (todos P > 0,05). En relación con el grupo imitador NC y el grupo si-NC, la expresión de VEGF y MMP-9 en el grupo imitador miR-133a-3p y el grupo si-PSAT1 se redujeron (todos P <0,05). Frente al grupo imitador de miR-133a-3p, la expresión de VEGF y MMP-9 se elevó en el grupo imitador de miR-133a-3p + oe-PSAT1 (todos P <0.05) (Fig. 7d, e).
PSAT1 es un gen objetivo de miR-133a-3p
Se utilizó el software de predicción en línea (https://cm.jefferson.edu/rna22/Precomputed/) para pronosticar y analizar el sitio objetivo de la unión de PSAT1 al miR-133a-3p, y la secuencia de la región 3'UTR combinada por PSAT1 y miR-133a-3p. Para demostrar que el sitio de unión predicho de miR-133a-3p daba como resultado un cambio en la actividad de luciferasa, se idearon la secuencia de mutación y la secuencia salvaje del sitio de unión de miR-133a-3p que suprime PSAT1 3'UTR. La actividad luciferasa se verificó mediante cotransfección de plásmidos recombinantes de miR-133a-3p y WT (Wt-miR-133a-3p / PSAT1) o MUT (Mut-miR-133a-3p / PSAT1) en EC vasculares. Los resultados revelaron que el imitador de miR-133a-3p no tuvo un efecto distintivo sobre la actividad de la luciferasa en el grupo Mut-miR-133a-3p / PSAT1 ( P > 0,05), mientras que la actividad luciferasa en el grupo Wt-miR-133a-3p / PSAT1 se redujo notablemente ( P <0.05) (Fig. 7f, g).
Discusión
IA es una dilatación anormal de la arteria intracraneal, que debilita la pared arterial al empujar continuamente hacia afuera la pared vascular, lo que da como resultado un mayor riesgo de rotura del aneurisma [17]. En un estudio realizado por Liu et al., Se ha demostrado que algunos miRNA están implicados en la modulación de la proliferación celular de las células del músculo liso vascular, que está estrechamente asociado con IA [18]. Además, un estudio reciente ha proporcionado una prueba de que los miARN circulantes pueden usarse como un nuevo biomarcador para evaluar la posibilidad de que ocurra IA en individuos de alto riesgo [19]. Se considera habitualmente que PSAT1 puede estar implicado en las condiciones del espectro de la esquizofrenia y altera el metabolismo de la serina [20]. El estudio actual fue diseñado para explorar el papel regulador de la lesión endotelial vascular modulada por miR-133a-3p y desencadenó IA mediante la modulación de la vía de señalización PSAT1 / GSK3β / β-catenina.
En el presente estudio, se analizó la relación entre la expresión de miR-133a-3p y las características clínico-patológicas de IA, y los resultados demostraron que la expresión de miR-133a-3p no estaba relacionada con la edad, el sexo, la forma y la posición del aneurisma, sino asociado con el número y tamaño del aneurisma. Algunos estudiosos consideraron que el esfuerzo cortante del flujo sanguíneo regional en la pared arterial indujo la inducción de la expresión de la proteína quimioatrayente de monocitos 1 (MCP-1) y la proteína inflamatoria de macrófagos 1α (MIP-1α) por los fibroblastos y las CE vasculares dentro de la pared vascular. Los factores quimiotácticos altamente reactivos MCP-1 y MIP-1α hicieron una agregación de macrofagocitos en la pared vascular y mediaron la respuesta inflamatoria, luego, indujeron la excitación del factor de transcripción nuclear c-Jun y luego regularon la activación de la proteína 1 activada (AP- 1), luego activó el promotor MMP-9 en su dominio estructural para aumentar la expresión del ARNm de MMP-9 y finalmente indujo la disolución de la matriz extracelular de la pared vascular, lo que provocó la formación de un aneurisma intracraneal [21,22,23]. Saito y col. [24] encontraron que las células positivas para MMP-9 provenían principalmente de las membranas media y externa de los macrófagos arteriales, lo que certificó que la MMP-9 expresada por macrófagos mediaba la degeneración de la pared arterial, lo que conducía a la formación de aneurisma arterial. Los estudios anteriores han indicado que MMP-9 estaba relacionada con la formación de IA. Los resultados de nuestro estudio revelaron que MMP-9 estaba regulado al alza en IA; por lo tanto, especulamos que mR-133a-3p podría estar involucrado en la aparición y desarrollo de IA regulando la vía PSAT1 / GSK3β / β-catenina y regulando adicionalmente MMP-9. En nuestro estudio, encontramos que la restauración de miR-133a-3p redujo la expresión de PSAT1, GSK3β, β-catenina y MMP-9 en tejidos de aneurismas intracraneales. Realizaremos investigaciones relevantes en el estudio futuro para verificar nuestros hallazgos.
Nuestro estudio ha proporcionado evidencia sustancial en relación con la noción de que la expresión de miR-133a-3p se redujo y el PSAT1, GSK3β y β-catenina se elevó en IA. La evidencia emergente ha sugerido que miR-133a-3p juega un papel supresor en diferentes tipos de tumores. Un estudio reciente ha presentado que la expresión de miR-133a-3p se degradó drásticamente en los tejidos del cáncer de mama en contraste con la de los tejidos no cancerosos [25]. Otro estudio ha pretendido que la expresión de miR-133a-3p está disminuida en los tejidos del cáncer de próstata avanzado (CaP) en relación con la de los tejidos normales adyacentes o los tejidos con lesiones benignas de la próstata, especialmente en los tejidos con CaP metastásico óseo [26]. El efecto promotor de PSAT1 en otros tipos de enfermedades se encuentra en algunas publicaciones. Se ha informado de que la expresión de PSAT1 aumentó notablemente en el cáncer de pulmón de células no pequeñas (CPCNP) y pronosticó un resultado clínico precario de los pacientes con CPCNP [27]. Además, PSAT1 se considera como el gen más elevado en los tumores de CCR, así como altamente expresado en pacientes con enfermedad quimiorresistente [28]. Se ha manifestado que la actividad de GSK3β estaba elevada en tejidos cancerosos [29]. Además, el nivel de fosforilación de GSK3β así como la expresión de la β-catenina nuclear también se mejoran, lo que sugiere que la vía de GSK3β / β-catenina puede participar en la regulación de la osteopontina [30].
Otros resultados surgieron de nuestros datos sugirieron que la regulación positiva de miR-133a-3p y la regulación negativa de PSAT1 suprimieron la apoptosis y la proliferación y migración avanzadas de las CE de IA, redujeron la expresión de VEGF y MMP-9 en tejidos de IA. Se ha sugerido anteriormente que la sobreexpresión de miR-133a-3p vuelve a entrenar la invasión, el crecimiento y la mitosis de las células del carcinoma de células escamosas orales al dirigirse al colágeno tipo I alfa 1 (COL1A1) [31]. Se ha informado de que miR-133a-3p altamente expresado puede reprimir la propagación de células ESCC, hacer avanzar la apoptosis celular y disminuir la migración e invasión de células ESCC al dirigirse a COL1A1 [32]. Otro estudio ha verificado que la regulación positiva transitoria de miR-133a-3p suprime la migración, la invasión y la capacidad de crecimiento de las células del carcinoma de vesícula biliar al dirigirse directamente a la proteína de unión a la señal de recombinación Jκ [33]. De la misma manera, este estudio sugiere que miR-133a-3p ejerce su papel en IA a través de la focalización en PSAT1. Se muestra que la sobreexpresión de PSAT1 estimula el crecimiento de células ESCC y la invasión de matrigel in vitro, y la inyección de ratones con células ESCC con alta expresión de PSAT1 induce la formación de tumores in vivo [14]. Otro estudio también ha informado de que PSAT1 está altamente expresado y pronostica un mal resultado clínico de los pacientes, así como mejora la tumorigénesis celular y la proliferación in vivo e in vitro [13]. Investigaciones previas generalmente confirman que PSAT1 promueve la progresión, proliferación y tumorigénesis del ciclo celular a través de experimentos de pérdida y ganancia de función [27]. Se ha indicado que las MMP están compuestas por una serie de enzimas que escinden los sustratos de proteínas sobre la base de un mecanismo conservado que se refiere a la activación de una molécula de agua unida al sitio activo a través de un Zn 2+ ion [34]. MMP-9 es una proteasa distinta que impulsa una inmensa influencia en muchos procesos biológicos [35]. Un estudio ha sostenido que la MMP-9 está elevada en los grupos de aneurismas en comparación con el grupo de control [36]. El factor de crecimiento del endotelio vascular A (VEGF-A) se reconoce como el modulador clave de la diferenciación endovascular del trofoblasto [37]. Un estudio ha revelado que la expresión de PSAT1 silenciada suprime la expresión de fosforilación de VEGF, β-catenina y GSK3β [10].
Conclusión
Para concluir brevemente, nuestro estudio confirma nuestra hipótesis de que la sobreexpresión de miR-133a-3p o la regulación a la baja de PSAT1 restringen el daño de las células endoteliales y avanzan la proliferación de células endoteliales mediante la inhibición de la vía GSK3β / β-catenina en IA. Estos hallazgos proporcionan una nueva perspectiva sobre una nueva terapia diana para la AI. Estos hallazgos subrayan el papel de miR-133a-3p en IA en relación con la vía PSAT1 / GSK3β / β-catenina. Sin embargo, no se puede llegar a una conclusión clara sobre los efectos de miR-133a-3p y PSAT1 debido a las limitadas investigaciones conocidas al respecto. Debe ser monitoreado rigurosamente e informado de manera apropiada en los ensayos clínicos futuros.
Disponibilidad de datos y materiales
No aplica
Abreviaturas
- miR-133a-3p:
-
MicroARN-133a-3p
- IA:
-
Aneurisma intracraneal
- PSAT1:
-
Fosfoserina aminotransferasa 1
- GSK3:
-
β-glucógeno sintasa quinasa 3β
- SAH:
-
Hemorragia subaracnoidea
- MiARN:
-
MicroARN
- CRC:
-
Cáncer colorrectal
- ESCC:
-
Carcinoma de células escamosas de esófago
- HE:
-
Hematoxilina-eosina
- PBS:
-
Solución salina tamponada con fosfato
- MMP:
-
Metaloproteasa de matriz
- VEGF:
-
Factor de crecimiento endotelial vascular
- NC:
-
Control negativo
- DAB:
-
Diaminobencidina
- FBS:
-
Suero fetal bovino
- UEAI:
-
Aglutininas I de Ulex europaeus
- PI:
-
Yoduro de propidio
- FITC:
-
Isotiocianato de V-fluoresceína
- DMSO:
-
Dimetilsulfóxido
- OD:
-
Densidad óptica
- SD:
-
Sprague-Dawley
- SPF:
-
Libre de patógenos específicos
- GAPDH:
-
Gliceraldehído fosfato deshidrogenasa
- BCA:
-
Ácido bicinconínico
- 3′UTR:
-
3 ′ Región sin traducir
- WT:
-
Tipo salvaje
- MUT:
-
Mutante
- ANOVA:
-
Análisis de varianza
- NSCLC:
-
Cáncer de pulmón de células no pequeñas
- VEGF-A:
-
Factor de crecimiento endotelial vascular A
Nanomateriales
- El amplificador de emisor común
- Preparación de nanopartículas de mPEG-ICA cargadas con ICA y su aplicación en el tratamiento del daño celular H9c2 inducido por LPS
- La administración de nanopartículas de artesunato mejora la eficacia antitumoral activando la apoptosis celular mediada por mitocondrias
- Control del crecimiento de nanocables de selenuro de indio de alta uniformidad (In2Se3) mediante el proceso de recocido térmico rápido a baja temperatura
- Síntesis fácil de óxido de estaño mesoporoso similar a un agujero de gusano a través del autoensamblaje inducido por evaporación y las propiedades mejoradas de detección de gas
- Diseño y ajuste de la función de trabajo del grafeno mediante tamaño, modificación, defectos y dopaje:un estudio de teoría del primer principio
- Una investigación sobre una célula solar de silicio cristalino con una capa de silicio negro en la parte posterior
- Ajuste del modo de movimiento deslizante de los nanotubos de carbono mediante grupos de hidroxilo
- Absorción de luz eficaz utilizando rejillas piramidales de doble cara para células solares de silicio de película delgada
- El efecto de apoptosis en las células de cáncer de hígado de nanopartículas de oro modificadas con ácido litocólico
- Diminutas nanopartículas de fluoruro de tierras raras activan el crecimiento de las células tumorales mediante interacciones polares eléctricas