


Biocompatibilidad de nanoportadores de liposomas en el oído interno de rata después de la administración intratimpánica
Resumen
Los nanoportadores de liposomas (LPN) son potencialmente el futuro de la terapia del oído interno debido a su alta capacidad de carga de fármacos y su absorción eficiente en el oído interno después de una administración intratimpánica mínimamente invasiva. Sin embargo, falta información sobre la biocompatibilidad de las LPN en el oído interno. El objetivo del presente estudio es documentar la biocompatibilidad de los NLP en el oído interno después del parto intratimpánico. Se administraron LPN con o sin ácido gadolinio-tetra-azaciclo-dodecano-tetra-acético (Gd-DOTA) a las ratas mediante inyección transtimpánica. La distribución de las LPN que contienen Gd-DOTA en el oído medio e interno se siguió in vivo mediante resonancia magnética. La función de las barreras del oído medio e interno se evaluó mediante resonancia magnética mejorada con gadolinio. La función auditiva se midió mediante la respuesta auditiva del tronco encefálico (ABR). La respuesta inflamatoria potencial se investigó analizando la secreción de glicosaminoglicano y ácido hialurónico y la expresión de CD44 y TLR2 en el oído interno. La apoptosis potencial se analizó utilizando transferasa terminal (TdT) para marcar las roturas libres 3'OH en las cadenas de ADN de células apoptóticas con TMR-dUTP (tinción TUNEL). Como resultado, los LPN ingresaron al oído interno de manera eficiente después de la inyección transtimpánica. La inyección transtimpánica de LPN con o sin Gd-DOTA no interrumpió la función de las barreras del oído medio e interno ni causó problemas de audición en ratas. Los marcadores biológicos inflamatorios críticos en el oído interno, incluida la secreción de glicosaminoglicanos y ácido hialurónico y la expresión de CD44 y TLR2, no se vieron influenciados por la administración de LPN. No hubo muerte celular significativa asociada con la administración de LPN. La inyección transtimpánica de LPN es segura para el oído interno, y las LPN se pueden aplicar como una matriz de administración de fármacos en la terapia clínica de la pérdida auditiva neurosensorial.
Antecedentes
Los nanoportadores de liposomas (LPN) son potencialmente el futuro de la terapia del oído interno debido a su alta capacidad de carga de fármacos y su absorción eficiente en el oído interno después de una administración intratimpánica mínimamente invasiva [1, 2, 3, 4]. El enfoque intratimpánico es bien aceptado por los otólogos como un enfoque racional de administración de fármacos dirigidos porque evita la acumulación innecesaria de agentes terapéuticos en regiones no dirigidas, lo que ha sido una estrategia previa en la clínica para el tratamiento de la enfermedad de Meniere y la pérdida auditiva neurosensorial súbita. usando gentamicina y corticosteroides. La dirección molecular de los modelos terapéuticos en la cóclea fue indicada por la administración intratimpánica de LPN funcionalizados con péptidos específicos [5]. Además, la administración sostenida automática de LPN al oído interno a través del oído medio se logró mediante un dispositivo novedoso compuesto por una bomba osmótica y un tubo de poliimida de alto rendimiento [6]. Como la plataforma nanoterapéutica más antigua de la clínica, las LPN eran seguras para tratar el cáncer, las enfermedades infecciosas, la inflamación, el dolor, etc. [7, 8, 9]. Sin embargo, la biocompatibilidad de los LPN en el oído medio e interno sigue siendo desconocida y debe aclararse antes de que puedan aplicarse clínicamente en otología.
El oído está compuesto por oídos externo, medio e interno (Fig. 1) que pueden estar expuestos a los LPN después del parto intratimpánico. El oído medio es el sitio principal que está expuesto a los LPN en su concentración más alta, el oído interno es el sitio terapéutico y el órgano más sensible a los agentes peligrosos, y el canal auditivo externo tiene el potencial de irritarse por los agentes que fluyen desde el cavidad del oído medio. Las barreras biológicas son el primer sistema de defensa, limitando la biodisponibilidad de los agentes y existen en la piel, las mucosas y las estructuras perineurales. El sistema de barrera en el oído interno juega un papel crítico en el mantenimiento de la homeostasis iónica que es esencial para la actividad fisiológica del oído interno. La alteración funcional de estas barreras se puede evaluar con precisión utilizando imágenes de resonancia magnética mejoradas con gadolinio (Gd-MRI). El deterioro de la función auditiva se puede medir con precisión a través de la respuesta auditiva del tronco encefálico (ABR). Por lo tanto, el oído (incluidos los oídos externo, medio e interno) en sí mismo sirve como un modelo excelente para la nanotoxicología [10, 11].
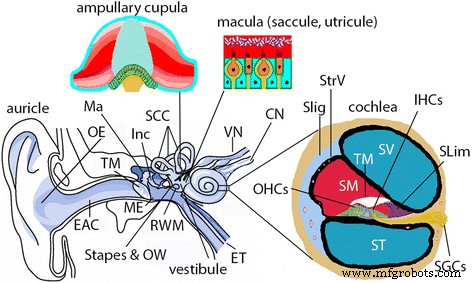
Ilustración de la oreja de mamífero. El oído de los mamíferos (incluidos los humanos y las ratas) se compone de los oídos externo, medio e interno. El oído externo ( OE ) se compone de aurícula y conducto auditivo externo ( EAC ). El oído medio ( YO ) está compuesto por la membrana timpánica ( TM ) y la cavidad que alberga la cadena osicular, incluido el martillo ( Ma ), incus ( Inc ) y estribo. La cavidad del oído medio es una extensión de la nasofaringe a través de la trompa de Eustaquio ( ET ) y se comunica con el oído interno a través de la ventana ovalada ( OW ) y membrana de ventana redonda ( RWM ). El oído interno está compuesto por cóclea y sistema vestibular. La cóclea es el órgano sensorial para la audición y tiene tres cámaras, es decir, los compartimentos perilinfáticos de la escala tímpano ( ST ) y scala vestibuli ( SV ) y el compartimento endolinfático de scala media ( SM ). En la pared lateral de SM, están las estrías vasculares ( StrV ) y ligamento espiral ( SLig ). En la parte inferior de SM, hay un órgano de Cortis que contiene células ciliadas internas ( IHC ) y células ciliadas externas ( OHC ), membrana tectorial ( TM ) y limbo en espiral ( Slim ). Las células ganglionares espirales ( SGC ) disparan un potencial de acción correspondiente a la transducción mecanoeléctrica de las células ciliadas y suministran toda la información auditiva del cerebro. El sistema vestibular es responsable del equilibrio y está compuesto por tres canales semicirculares ( SCC ) y vestíbulo. La cúpula ampular dentro del SCC detecta aceleraciones rotacionales y la mácula dentro del sáculo y utrículo del vestíbulo detecta aceleraciones lineales. CN nervio coclear, SP prominencia en espiral, VN nervio vestibular, VS vas spiralis. (adaptado de Zou J. Focal Drug Delivery in Inner Ear Therapy:in Focal Controlled Drug Delivery. Editores:Domb AJ y Khan W. Springer, Londres, Reino Unido. ISBN:978-1-4614-9433-1, 2014; p215- 224)
El ácido hialurónico (hialuronano) es un biopolímero polianiónico de origen natural y es un componente principal de la matriz extracelular en la membrana basal. El ácido hialurónico está compuesto por ácido D-glucurónico y N-acetil-D-glucosamina, que están unidos mediante enlaces glucosídicos β-1, 4 y β-1, 3 alternos. La acumulación de ácido hialurónico podría contribuir al aumento de la permeabilidad y la inflamación de la microcirculación en la lesión por reperfusión isquémica renal [12]. En nuestro informe anterior se demostró que el efecto ototóxico de las nanopartículas de plata se correlaciona con la acumulación de ácido hialurónico en la cóclea de rata [11]. El ácido hialurónico se une al CD44 y al receptor tipo toll 2/4 (TLR2 / 4) en el tejido y desencadena reacciones biológicas [13, 14]. Las actividades biológicas mediadas por CD44 al unirse al ácido hialurónico son principalmente a través de la interacción con moléculas reguladoras y adaptadoras, como SRC quinasas, Rho GTPasas, VAV2, proteína de unión asociada a proteína 2 unida al receptor del factor de crecimiento 1 (GAB1), anquirina, y ezrin [15, 16, 17]. CD44 también media en el metabolismo del ácido hialurónico a través de enfoques de captación y degradación celular, además de reclutar células T en sitios inflamatorios y regular la lesión endotelial mediada por células T [18]. Se informó que la citotoxicidad de las células endoteliales del oído interno por los anticuerpos anti-células endoteliales podría desempeñar un papel en la causa del daño de la estría vascular en la sordera neurosensorial súbita inmunomediada [19]. Se informó que la activación del factor nuclear κB dependiente de TLR2 estuvo involucrada en la regulación positiva de la proteína 1 quimiotáctica de monocitos inducida por Haemophilus influenzae no tipificable en los fibrocitos del ligamento espiral del oído interno, que podría ser el paso clave en la disfunción del oído interno secundaria a otitis media crónica [ 20]. Si los LPN inducen deterioro del oído interno después de la administración en el oído medio, la vía de señalización mediada por TLR2 debería ser el mecanismo importante.
Nuestro objetivo fue evaluar la biocompatibilidad de LPN en el oído interno después de la inyección transtimpánica. Las funciones de las barreras biológicas en la piel (canal auditivo externo), mucosa (cavidad del oído medio) y compartimentos del oído interno se midieron utilizando Gd-MRI en varios momentos. La función auditiva se evaluó mediante la medición de ABR. Finalmente, se analizaron los posibles cambios histopatológicos midiendo las acumulaciones de glicosaminoglicanos y ácido hialurónico, las expresiones de CD44 y TLR2 y la fragmentación del ADN en la cóclea.
Resultados
Los LPN no causaron cambios funcionales en la cóclea de rata
En el grupo de control positivo, señal brillante en la perilinfa de la cóclea (Coch) y vestibular (Vest) en ambos lados (L, R) (Fig. 2a, b) que indica captación de Gd-DOTA. Después de las inyecciones transtimpánicas de nanopartículas de plata (AgNP), las intensidades de la señal en los compartimentos perilinfáticos aumentaron significativamente, mientras que también se detectó una señal extremadamente intensa en la piel del conducto auditivo externo, la mucosa del oído medio, lo que indica una mayor captación de Gd-DOTA asociada con la administración de AgNP ( L en la Fig. 2a, b) (Tabla 1). Por tanto, se validó el sistema de evaluación. En el animal que recibió una inyección transtimpánica de LPN + Gd-DOTA, se detectó una señal brillante en la superficie de la cadena osicular, el vestíbulo de la escala, la escala del tímpano y el vestíbulo 3 h después de la inyección, lo que indica una distribución obvia de la LPN en estas regiones (Fig. 2c). , D). La intensidad de la señal en el scala vestibuli en el giro basal fue visiblemente más fuerte que en el scala tympani sugiriendo una entrada eficiente de LPN a través de la ventana oval en el animal actual [21]. A las 6 h después de la inyección, las intensidades de señal entre la escala vestibular y la escala timpánica en el giro basal se volvieron similares y toda la cóclea mostró una señal casi homogénea, pero hubo cambios insignificantes en el vestíbulo (Fig. 2e, f). En los animales que recibieron inyecciones intravenosas de Gd-DOTA después de la inyección transtimpánica de LPN en blanco, ambos lados mostraron intensidades de señal similares, excepto que hubo señales fuertes en el oído medio que recibieron una inyección transtimpánica de LPN en blanco que sospechaba acumulación de LPN en la superficie de la cadena osicular (Fig. .2g, h). Agujero negro en la cadena de huesecillos que indica la zona hueca del estribo (fig. 2h). Intensidades de señal iguales en ambos lados sugirieron que la propiedad de transporte para Gd-DOTA de las barreras sangre-perilinfa en ambos oídos no cambió después de la inyección transtimpánica de LPN (Fig. 2g, h) (Tabla 1).
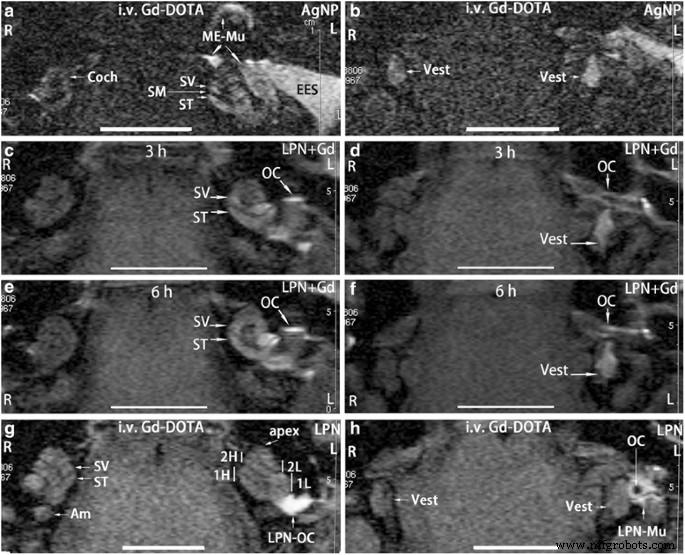
Resonancia magnética mejorada con gadolinio del oído interno de rata después de la administración de nanoportadores de liposomas (LPN). En todos los animales, se inyectaron nanomateriales en la pared medial de la cavidad del oído medio izquierdo. Se obtuvieron imágenes del control positivo en ratas 2 h después de la inyección intravenosa de Gd-DOTA secundaria a la inyección transtimpánica de nanopartículas de plata (AgNP) 5 h antes ( a , b ). La distribución dinámica de LPN en el oído medio e interno se muestra en c , d , e , f mediante inyección transtimpánica de LPN que contiene Gd-DOTA sin administración intravenosa de Gd-DOTA. El impacto de los LPN vacíos sobre la barrera biológica se demostró mediante resonancia magnética 2 h después de la inyección intravenosa de Gd-DOTA (i.v. Gd-DOTA) en ratas que recibieron una inyección transtimpánica de LPN 5 h antes ( g , h ). Soy ampular del canal semicircular posterior, Coch cóclea, EES piel del oído externo, L oreja izquierda, LPN-Mu LPN en la mucosa del oído medio, LPN-OC LPN en la cadena osicular ( OC ), ME-Mu , mucosa del oído medio, R oído derecho, SM scala media , ST scala tympani, SV scala vestibuli, chaleco vestibulum, 1H giro basal superior de la cóclea, 1L vuelta basal inferior de la cóclea, 2H segunda vuelta más alta de la cóclea, 2L segunda vuelta inferior de la cóclea. Barra de escala =5 mm
Ni LPN + Gd-DOTA ni LPN causaron una pérdida auditiva significativa, presentada como un cambio de umbral ABR que se midió utilizando estímulos de clic y ráfagas de tono en las frecuencias de 2, 4, 8, 16 y 32 kHz a 2, 4 y 7 días después de la administración, en comparación con los oídos que recibieron inyecciones transtimpánicas de agua desionizada (dH 2 O) (Figura 3).
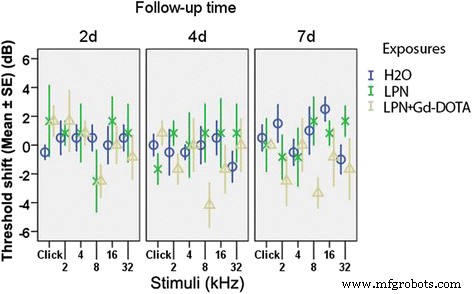
Impacto de la inyección transtimpánica de nanoportadores de liposomas en la función auditiva en ratas medida por la respuesta auditiva del tronco encefálico. La pérdida auditiva se expresó como cambios de umbral. Hubo una diferencia insignificante entre los grupos ( p > 0.05, ANOVA unidireccional). n =6 en cada grupo. H2O inyección transtimpánica de agua desionizada en el grupo de control negativo, LPN nanoportador de liposomas vacío, LPN + Gd-DOTA LPN que contiene Gd-DOTA, 2d , 4d , y 7d 2, 4 y 7 días después de la inyección
Los LPN no indujeron la acumulación de glucosaminoglicanos en la cóclea de rata
La tinción con hematoxilina y eosina no demostró ninguna infiltración inflamatoria de leucocitos y fibrina en la cóclea de todos los animales analizados, incluido el estribo y la ventana oval por donde pasan los LPN (Fig. 4). La tinción con ácido periódico de Schiff demostró la existencia de glicosaminoglicanos en la pared ósea, limbo espiral, ligamento espiral, membrana tectorial, membrana de Reissner, lámina espiral ósea y estría vascular en la cóclea de animales que recibieron inyecciones transtimpánicas de dH 2 O. Hubo un aumento gradiente en la intensidad de la señal desde el giro basal hasta el ápice, y la diferencia fue significativa en la estría vascular (Figs. 5 y 6). El gradiente de señal en la cóclea no cambió en los animales que recibieron inyección transtimpánica de LPN y LPN + Gd-DOTA (Figuras 5 y 6).
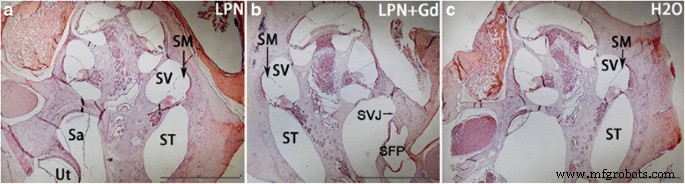
Tinción con hematoxilina-eosina de cócleas de rata expuestas a nanoportadores de liposomas. No hubo infiltración inflamatoria en la cóclea recibió administraciones de LPN ( a ), LPN + Gd ( b ) y H2O ( c ). Área rodeada selección indicada de la región de interés para las mediciones de intensidad ( a ). LPN nanoportador de liposomas vacío, LPN + Gd LPN que contiene Gd-DOTA. Sa sáculo, SFP plataforma del estribo, SVJ articulación estapediovestibular, SM scala media, ST scala tympani, SV scala vestibuli, Ut utricule. Barra de escala =1 mm
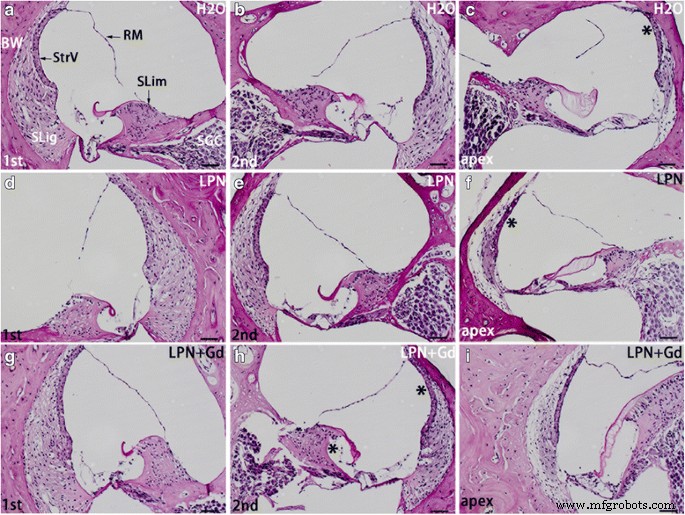
La secreción de glucosaminoglicanos en la cóclea de rata expuesta a nanoportadores de liposomas se detectó utilizando microscopía óptica de tinción de ácido periódico de Schiff. El limbo espiral ( SLim ) y pared ósea ( BW ) de la cóclea mostró la tinción más intensa en los grupos de control negativo ( H2O ) ( a - c ), nanoportador de liposomas vacío (LPN) ( d - f ) y LPN que contiene Gd-DOTA ( LPN + Gd ) ( g - yo ). El área de tinción con intensidades visiblemente más altas se indica con * en c , f y h en comparación con la columna de la izquierda. RM Membrana de Reissner, SGC célula ganglionar espiral, SLig ligamento espiral, StrV estría vascular, 1.ª giro basal, 2.º segundo turno. Barra de escala =50 micras
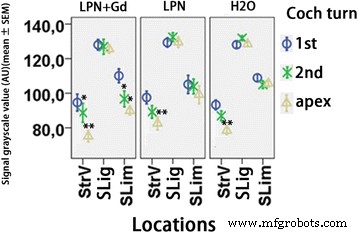
Cuantificación de la secreción de glicosaminoglicanos en cócleas de ratas expuestas a nanoportadores de liposomas detectadas mediante tinción de ácido periódico de Schiff. n =3 en cada grupo. AU unidad arbitraria, H2O control negativo, LPN nanoportador de liposomas vacío, LPN + Gd LPN que contiene Gd-DOTA, SLig ligamento espiral, delgado limbo espiral, StrV estría vascular, 1.ª giro basal, 2.º segundo turno. * p <0.05, ** p <0.01 (ANOVA unidireccional con prueba LSD utilizada como análisis post hoc)
Hubo un impacto menor en la secreción de ácido hialurónico en la cóclea de rata por LPNs
En la cóclea de ratas que recibieron inyecciones transtimpánicas de dH2O, se detectó tinción positiva para ácido hialurónico predominantemente en las células ganglionares espirales, células basales del estriado, células del surco externo y células endoteliales capilares, entre otras células (Fig. 7). Las intensidades de señal en los fibrocitos del ligamento espiral de las vueltas basal y segunda fueron significativamente más altas que las del ápice. Estas diferencias se volvieron insignificantes en la cóclea de ratas con la aplicación de LPN y LPN + Gd-DOTA, lo que indica que la secreción de ácido hialurónico por los fibrocitos del ligamento espiral se vio afectada por la administración de LPN (Fig. 8). LPN + Gd-DOTA también redujo la tinción en los fibrocitos del ligamento espiral del giro basal. Sin embargo, no hubo impacto en la secreción de ácido hialurónico en la mayoría de las células cocleares por la inyección transtimpánica de LPN y LPN + Gd-DOTA (Figuras 7 y 8).
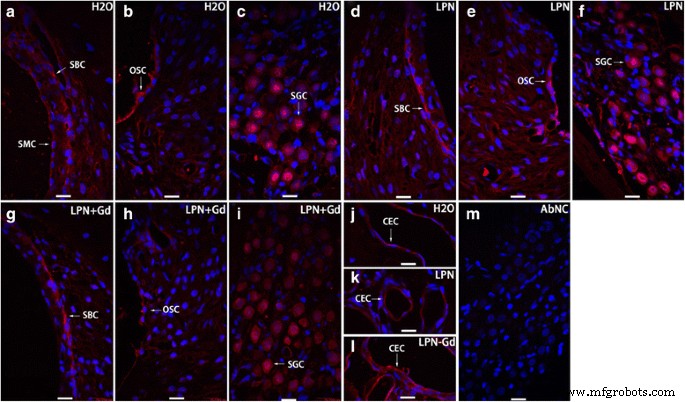
La secreción de ácido hialurónico en la cóclea de rata expuesta a nanoportadores de liposomas se detectó con microscopía confocal inmunofluorescente. Se encontró tinción positiva en las células basales de la estría ( SBC ), celda del surco externo ( OSC ), célula ganglionar espiral ( SGC ) y células endoteliales capilares ( CEC ) de modiolus de grupos de control negativo ( H2O ) ( a - c , j ), nanoportadores de liposomas vacíos ( LPN ) ( d - f , k ) y LPN que contiene Gd-DOTA ( LPN + Gd ) ( g - yo , l ). No hubo tinción en el anticuerpo omitido control negativo ( AbNC ) ( m ). CEC célula endotelial capilar, ISC celda del surco interno, SBC célula basal estriada, SL-I fibrocito del ligamento espiral tipo I, SLSF fibrocito satélite del limbo en espiral, SMC stria vascularis célula marginal. Barra de escala =16 micras
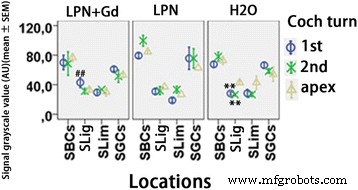
Cuantificación de la secreción de ácido hialurónico en cócleas de ratas expuestas a nanoportadores de liposomas detectadas mediante microscopía confocal inmunofluorescente. n =3 en cada grupo. AU unidad arbitraria, H2O control negativo, LPN nanoportador de liposomas vacío, LPN + Gd SBC que contienen Gd-DOTA células basales estriadas, SGC células ganglionares espirales, SLig , ligamento espiral, delgado limbo espiral, 1. ° giro basal, 2.º segundo turno. ** p <0.01 (en comparación con el ápice), ## p <0.01 (en comparación con los grupos LPN y H2O del giro basal) (ANOVA unidireccional con prueba LSD utilizada como análisis post hoc)
Los LPN no alteraron la población de células CD44 en la cóclea de rata
En las cócleas expuestas a dH 2 O, las células intermedias del estriado, las células basales del estriado, los fibrocitos del ligamento espiral, las células del ganglio espiral, las células de Deiters en el órgano de Corti y las células del endotelio capilar en el modiolo y el ligamento espiral mostraron una tinción intensa para CD44. Hubo una diferencia insignificante en las intensidades de la señal entre los giros cocleares. La población positiva para CD44 y la intensidad de expresión no se vieron afectadas por la inyección transtimpánica de LPN + Gd-DOTA o LPN (Figuras 9 y 10).
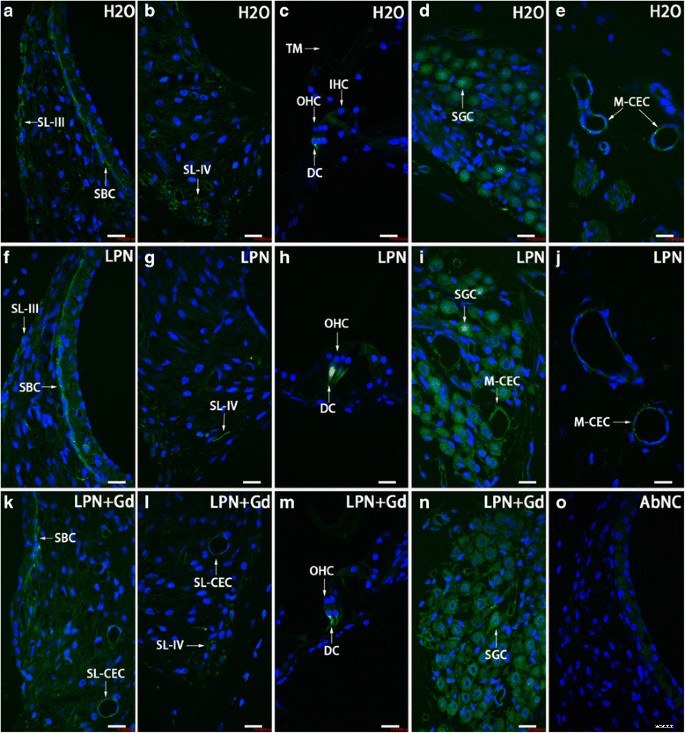
Distribución de células CD44-positivas en la cóclea de rata expuesta a nanoportador de liposomas demostrada usando microscopía confocal inmunofluorescente. Las células CD44 positivas se detectaron principalmente en las células basales de la estría ( SBC ), ligamento espiral ( SL ), Células de Dieter ( DC ), célula ganglionar espiral ( SGC ) y células endoteliales capilares ( CEC ) en los grupos de control negativo ( H2O ) ( a - e ), nanoportadores de liposomas vacíos ( LPN ) ( f - j ) y LPN que contiene Gd-DOTA ( LPN + Gd ) ( k - n ). No hubo tinción en el anticuerpo omitido control negativo ( AbNC ) ( o ). IHC células ciliadas internas, M-CEC célula endotelial capilar en modiolo, SL-CEC célula endotelial capilar en ligamento espiral:células ganglionares espirales, SL-III fibrocito del ligamento espiral tipo III, SL-IV fibrocito del ligamento espiral tipo IV, TM membrana tectorial, OHC células ciliadas externas. Barra de escala =16 micras
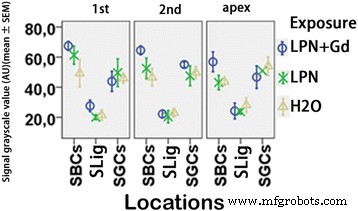
Cuantificación del nivel de proteína CD44 en cóclea de rata expuesta a nanoportadores de liposomas detectada mediante microscopía confocal inmunofluorescente. Hubo una diferencia insignificante entre los grupos ( p > 0.05, ANOVA unidireccional). n =3 en cada grupo. n =3 en cada grupo. AU unidad arbitraria, H2O control negativo, LPN nanoportador de liposomas vacío, LPN + Gd SBC que contienen Gd-DOTA células basales estriadas, SGC células ganglionares espirales, SLig ligamento espiral, 1.º giro basal, 2.º segundo turno
Los LPN no alteraron la expresión de TLR2 en la cóclea de rata
En las cócleas expuestas a dH2O, las células basales del estriado, los fibrocitos del ligamento espiral, las células de la raíz, las células del ganglio espiral, las células pilares del órgano de Corti y las células endoteliales capilares del modiolo mostraron una tinción intensa para TLR2. Hubo una diferencia insignificante en las intensidades de la señal entre los giros cocleares. La población TLR2 positiva y la intensidad de expresión no se vieron afectadas por la inyección transtimpánica de LPN + Gd-DOTA o LPN (Figuras 11 y 12).
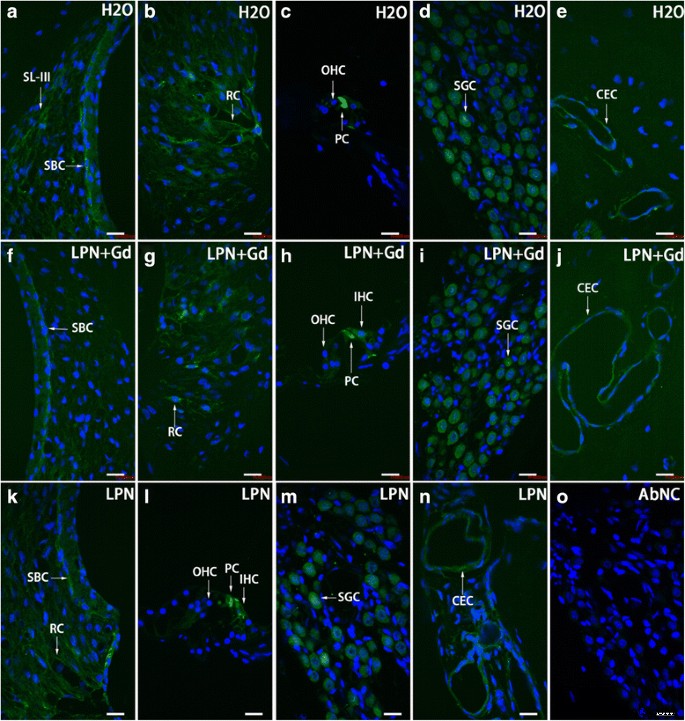
Distribución de células TLR2-positivas en la cóclea de rata expuesta a nanoportadores de liposomas demostrada usando microscopía confocal inmunofluorescente. Las células TLR2 positivas se detectaron principalmente en las células basales de la estría ( SBC ), ligamento espiral ( SL ), celda raíz ( RC ), celda de pilar ( PC ), célula ganglionar espiral ( SGC ) y células endoteliales capilares ( CEC ) en los grupos de control negativo ( H2O ) ( a - e ), nanoportadores de liposomas vacíos ( LPN ) ( k - n ) y LPN que contiene Gd-DOTA ( LPN + Gd ) ( f - j ). No hubo tinción en el control negativo omitido de anticuerpos ( AbNC ) ( o ). IHC células ciliadas internas, SL-III fibrocito de ligamento espiral tipo III, OHC células ciliadas externas. Barra de escala =16 micras
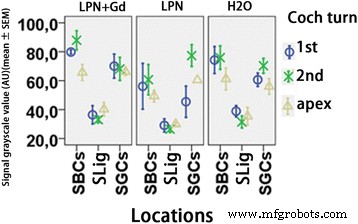
Cuantificación del nivel de proteína TLR2 en cócleas de ratas expuestas a nanoportadores de liposomas detectadas mediante microscopía confocal inmunofluorescente. Hubo una diferencia insignificante entre los grupos ( p > 0.05, ANOVA unidireccional). n =3 en cada grupo. AU unidad arbitraria, H2O control negativo, LPN nanoportador de liposomas vacío, LPN + Gd SBC que contienen Gd-DOTA células basales estriadas, SGC células ganglionares espirales, SLig ligamento espiral, 1.º giro basal, 2.º segundo turno
Los LPN no causaron la muerte celular en la cóclea de rata
Había escasas células apoptóticas que se distribuían al azar en la cóclea de ratas no tratadas. Sorprendentemente, había abundantes células apoptóticas en la base del estribo y el nicho de la ventana oval. No hubo impacto en la cantidad y el patrón de distribución de células apoptóticas por la administración de LPN y LPN + Gd-DOTA (Fig. 13).
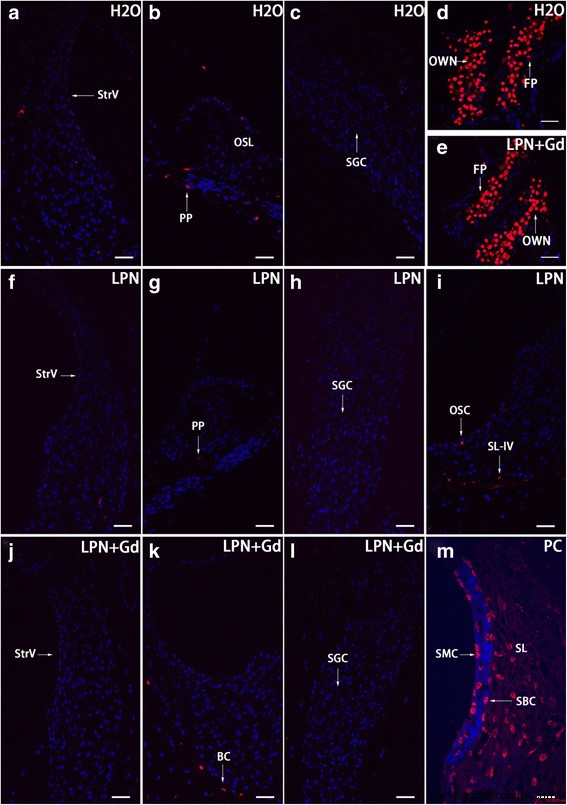
Apoptosis en la cóclea de rata expuesta a nanoportadores de liposomas demostrada usando microscopía confocal de tinción TUNEL. Se detectaron escasamente células apoptóticas en las células cocleares de ratas en grupos de control negativo ( H2O ) ( a - c ), nanoportadores de liposomas vacíos ( LPN ) ( f - yo ) y LPN que contiene Gd-DOTA ( LPN + Gd ) ( j - l ). Había abundantes células apoptóticas en la base del estribo ( FP ) y nicho de ventana ovalada ( PROPIO ) de ambos grupos ( d , e ). En un control positivo (PC), se detectó abundante tinción TUNEL en los fibrocitos del ligamento espiral ( SL ), células basales estriadas ( SBC ) y células de la estría marginal ( SMC ). PP proceso periférico de la célula ganglionar espiral ( SGC ), OC osteocito, OSC celda del surco externo, SL-IV tipo IV de fibrocitos del ligamento espiral, SLim limbo espiral, StrV stria vascularis. Barra de escala un - l =32 μm, m =16 micras
Discusión
Los LPN ingresaron al oído interno de manera eficiente después de la inyección transtimpánica demostrada por resonancia magnética utilizando Gd-DOTA como fármacos miméticos que se encapsularon dentro de los LPN (fig. 2c-f). Aunque un estudio anterior mostró que la ventana redonda era la vía principal de los LPN para ingresar al interior [6], la presente observación mostró que la vía de la ventana oval era más eficiente que la ventana redonda para transportar los LPN desde el oído medio hacia el interior. oreja. Este resultado sugirió que ambas vías son importantes en la carga de LPN en el oído interno después de la administración dirigida a la pared medial del oído medio. Utilizando el método in vivo más eficiente de resonancia magnética del oído interno con gadolinio para evaluar la barrera biológica y el ABR específico de frecuencia para evaluar la función auditiva, el presente estudio demostró que la inyección transtimpánica de LPN y LPN + Gd-DOTA no interrumpió la función de las barreras del oído interno ni causaron discapacidad auditiva en ratas. Al analizar los marcadores biológicos inflamatorios críticos previamente demostrados [11, 22], LPN y LPN + Gd-DOTA no indujeron la respuesta inflamatoria en la cóclea. Aunque no se evaluó la membrana de la ventana redonda, la ausencia de inflamación en el estribo y la ventana oval descartó una reacción inflamatoria obvia en la membrana de la ventana redonda, ya que el presente estudio demostró que la vía de la ventana oval era superior al abordaje de la ventana redonda para las NPL.
La resonancia magnética del oído interno después de la inyección intravenosa de quelato de gadolinio es capaz de detectar la alteración mediada por el estrés oxidativo en las barreras sangre-perilinfa y sangre-endolinfa inducida por toxinas mitocondriales [23]. Se informó que los AgNP causan deterioro celular a través de la generación de especies reactivas de oxígeno (ROS) y la activación de las quinasas amino-terminales Jun (JNK), lo que lleva a la liberación de citocromo C en el citosol y la translocación de Bax a las mitocondrias [24 ]. En la inyección transtimpánica, los AgNP entraron en el oído interno e indujeron cambios de permeabilidad en las barreras biológicas del oído interno de la rata [11, 25]. Los LPN también ingresaron al oído interno de la rata después de la inyección transtimpánica en un patrón dependiente del tamaño, y los LPN de 95 nm de diámetro mostraron la mayor eficacia para atravesar las barreras del oído medio-interno [3]. En el presente estudio, el tamaño medio de los LPN fue de 100 a 115 nm, que fue un poco más grande que el tamaño más eficiente. LPN + Gd-DOTA mostró que este tamaño de LPN ingresó al oído interno, lo que está de acuerdo con el informe anterior [3]. Sin embargo, la entrada de LPN en el oído interno no provocó cambios de permeabilidad en las barreras sangre-perilinfa y sangre-endolinfa. Este resultado sugirió que las LPN son seguras para el oído interno. Los resultados de ABR que indican una función auditiva normal respaldaron el resultado de la resonancia magnética.
There was an association between hyaluronic acid secretion and permeability change and microcirculation inflammation in renal ischemic reperfusion injury [12]. The previous study also showed that AgNPs caused the accumulation of hyaluronic acid in the rat cochlea [11]. CD44 and toll-like receptor 2/4 (TLR2/4) work as receptors of hyaluronic acid and trigger biological reactions [13, 14]. CD44 also mediates the metabolism of hyaluronic acid through cellular uptake and degradation in addition to recruiting T cells to inflammatory sites and regulating T cell-mediated endothelial injury [18]. In the present study, hyaluronic acid, CD44, and TLR2 were detected in the rat cochlea. LPN + Gd-DOTA reduced the secretion of hyaluronic acid in the spiral ligament fibrocytes. The expressions of CD44 and TLR2 were not changed after the transtympanic injection of either LPNs or LPN + Gd-DOTA. The total glycosaminoglycan, which contains hyaluronic acid in the cochlea was not affected by the administrations of LPNs and LPN + Gd-DOTA. The impact of LPNs on the hyaluronic acid distribution in rat cochlea did not cause either permeability change or hearing loss, indicating that the modification is unharmful. It was reported that macrophages undergo phenotypic changes dependent on molecular weight of hyaluronan that correspond to either pro-inflammatory response for low molecular weight hyaluronic acid or anti-inflammatory response for high molecular weight hyaluronic acid [26]. The observed minor changes of hyaluronic acid distribution in the cochlea might have high molecular weight and anti-inflammatory function. Therefore, there was no hint of an inflammatory reaction in the rat cochlea.
The observed apoptosis in the stapes footplate cells might be normal biological activity. A balance between survival and apoptosis in the stapes footplate cells was reportedly as necessary to inactivate the otosclerosis [27]. Administration of LPN + Gd-DOTA did not affect apoptosis in the rat stapes.
Conclusiones
The present study demonstrated that the transtympanic injection of liposome nanocarriers neither impaired the biological barriers of the inner ear nor caused hearing loss in the rats. The critical inflammatory mechanism was not activated by the administration of liposome nanocarriers, either. The results suggested that transtympanic injection of liposome nanocarrier is safe for the cochlea of rat.
Methods
Materials
Sphingosine (Sph), 1-stearoyl-2-oleoyl-sn-glycero-3-phosphocholine (SOPC), and 1, 2-distearoyl-sn-glycero-3-phosphoethanolamine-N-[methoxy(polyethyleneglycol)-2000] (ammonium salt) [DSPE-PEG-2000] were purchased from Avanti polar lipids (Alabaster, USA). DiI (Vybrant DiI cell-labeling solution, 1 mM in solvent) and N-(6-tetramethylrhodaminethiocarbamoyl)-1,2-dihexadecanoyl-sn-glycero-3-hosphoethanolamine, triethylammonium salt (TRITC-DHPE) were purchased from Thermo Fisher Scientific (Waltham, USA). Gd-DOTA (DOTAREM) was from Guerbet, Cedex, France. Hepes and EDTA were from Sigma. The purity of the lipids was evaluated using thin-layer chromatography on silicic acid-coated plates (Merck, Darmstadt, Germany) developed with a chloroform/methanol/water mixture (65:25:4, v/v/v). An examination of the plates after iodine staining and, when appropriate, upon UV illumination revealed no impurities. The lipid concentrations were determined gravimetrically with SuperG (Kibron, Espoo, Finland); a high-precision microbalance. The polyvinylpyrrolidone-stabilized AgNPs were supplied by Colorobbia (Firenze, Italy). The AgNPs were dispersed in deionized water (370.7 mM), and scanning electron microscopy showed that the AgNPs are spheroids with a particle size of around 100 nm. Dynamic light scattering (DLS) showed a mean hydrodynamic size of 117 ± 24 nm and a mean zeta potential of −20 ± 9 mV.
In the visualization of nanocarrier uptake in the inner ear and the evaluation of biological barrier function, 5 male Sprague Dawley rats, weighing between 334 and 348 g, were provided by the Biomedicum Helsinki, Laboratory Animal Centre, University of Helsinki, Finland (this is the defined animal center that provides animals for MRI experiments in Biomedicum); in the ABR and histological studies, 18 Sprague Dawley rats, weighing between 300 and 400 g, were provided by the Experimental Animal Unit of the University of Tampere School of Medicine in Finland. Animal assignments in each study were shown in Table 2. All animal experiments were approved by the Ethical Committee of University of Tampere (permission number:ESAVI/3033/04.10.03/2011). Animal care and experimental procedures were conducted in accordance with European legislation. Animals in the Gd-MRI study were anesthetized with isoflurane with 5% isoflurane–oxygen mixture (flow-rate 1.0 L/min) for induction and 3% for maintenance via a facemask. Animals for the ABR and histological studies were anesthetized with a mixture of 0.5 mg/kg medetomidine hydrochloride (Domitor ® , Orion, Espoo, Finland) and 75 mg/kg ketamine hydrochloride (Ketalar ® , Pfizer, Helsinki, Finland) via intraperitoneal injection followed by an intramuscular injection of enrofloxacin (Baytril ® vet, Orion, Turku, Finland) at a dose of 10 mg/kg to prevent potential infection. The animal’s eyes were protected by Viscotears® (Novartis Healthcare A/S, Copenhagen, Denmark).
Preparation and Characterization of LPNs with and without Gd
Preparation of Gd-containing LPNs
LPNs of unilamellar vesicles with an apparent hydrodynamic particle diameter (Z av ) of 110 ± 15 nm that contain Gd-DOTA were prepared according to the previously published method [6]. A concentration of 1 mM Gd-DOTA-containing LPN (LPN + Gd-DOTA) refers to 1 mM liposomes encapsulating 500 mmol/L of Gd-DOTA.
Preparation of Blank LPNs
Blank LPNs of unilamellar vesicles with Z av of 115 ± 10 nm were prepared according to a previous publication [6].
Administration of LPNs
Under general anesthesia with isoflurane with 5% isoflurane–oxygen mixture (flow-rate 1.0 L/min), 50 μl of either LPNs or LPN + Gd-DOTA were injected into the left middle ear cavity through the tympanic membrane penetration under an operating microscope (OPMI1-F, Carl Zeiss, Jena, Germany). The same amount of deionized water (H2 O) was injected transtympanically in rats that were assigned to the negative group. After the injection, the animals were kept in the lateral position with the injected ear oriented upward for 15 min before further measurements.
Evaluation on Biological Barrier Function Using Gd-MRI
One animal receiving transtympanic injection of LPN + Gd-DOTA was selected to demonstrate distributions of LPN in the inner ear using MRI without contrast agent. Two animals receiving transtympanic injection of blank LPNs were engaged in MRI study for evaluation of the biological barrier function. Two animals receiving transtympanic injection of AgNPs (370.7 mM, 40 μl) were used as positive control. The contralateral ear without any injection was used as negative control in all studies. A 4.7T MR scanner with a bore diameter of 155 mm (PharmaScan, Bruker BioSpin, Ettlingen, Germany) was utilized. The maximum gradient strength was 300 mT/m with an 80-μs rise time. A gadolinium-tetraazacyclododecane-tetraacetic acid (Gd-DOTA, 500 mM, DOTAREM, Guerbet, Cedex, France) solution was injected into the tail vein (0.725 mM/kg) 2 h before the MRI measurements. The imaging protocol and rapid acquisition with relaxation enhancement (RARE) sequences were applied according to a previous publication [10]. MRI scanning commenced at several time points after the transtympanic injection. The first MRI time of around 5 h was determined by taking the penetration time of liposome nanoparticles from the middle ear to the inner ear as a reference [1, 3, 6]. The final imaging time of 8 d was selected according to the course of potential acute inflammation and the availability of the scanner. ParaVision PV 4.0 (Bruker, MA, USA) software was used for the post-processing and quantification of MR images.
ABR Measurement
The auditory function of animals receiving injections of both blank and Gd-containing LPNs were evaluated using ABR measurements using BioSig32 (Tucker Davis Technologies, FL, USA) in a custom made, soundproof chamber. The ABR thresholds upon click and tone burst stimuli were recorded before and at a certain time point post-administration of LPNs. The first ABR measurement was followed on 2 days post-administration of AgNPs, allowing the animals to recover from the general anesthesia during the injection and to ensure the injected solution to be entirely cleared from the middle ear cavity. The second follow-up time of 4 days post-injection was chosen because it is close to the peak time of potential mitochondrial impairment-induced cell death in the cochlea [22]. The third follow-up time of 7 days is the time point when temporary threshold shifts remained significantly approved in an animal model of mitochondrial toxin-induced hearing loss [28]. The ABR recording procedure followed the previous report [11].
Glycosaminoglycan Staining in Rat Cochlea After Administration of LPNs
Hematoxylin-eosin staining to assess potential inflammatory infiltration and periodic acid Schiff’s staining to evaluate potential glycosaminoglycan accumulation in the cochlea after administration of LPNs were performed according to a previous publication after ABR measurements over 7 days [11] The slices were observed and digital images were acquired under a light microscope (Leica DM2000 microscope equipped with an Olympus DP25 camera) for further analysis.
Immunofluorescence Staining for Hyaluronic Acid and Receptors
Immunofluorescence staining for hyaluronic acid, CD44, and TLR2 were performed according to a previous publication after ABR measurements over 7 days [11, 21].
Cell Death Detection
Potential nuclear DNA fragmentation in the cochlea was investigated using terminal transferase (TdT) to label the free 3′OH breaks in the DNA strands of apoptotic cells with TMR-dUTP (TUNEL staining) following the reported procedure [11]. Slices exposed to recombinant DNase I (Fermentas, Vantaa, Finland, 100 U/ml in 50 mM Tris/HCl, pH 7.5, 1 mg/ml bovine serum albumin) at 37 °C for 10 min, which induced DNA strand breaks prior to the labeling procedures, were utilized as positive controls. The samples were observed under a confocal microscope.
Confocal Microscopy
The samples were observed under a Nikon inverted microscope (ECLIPSE Ti) combined with an Andor confocal system installed with Andor iQ 2.8 software (Andor Technology, Belfast, UK). The excitation lasers were 488 nm (green excitation) and 568 nm (red excitation) from an Andor laser combiner system, and the corresponding emission filters were 525/50 (Alexa Fluor-488) and 607/45 nm (Cy TM 3 and TMR Red). DAPI was excited with light at 405 nm generated from a light-emitting diode and was detected using a 450–465 nm filter.
Analysis and Statistics
ImageJ (1.45S, National Institutes of Health, Bethesda, USA) software was used for signal intensity measurements. For light microscopy of periodic acid Schiff’s staining, the region of interests (ROIs) including spiral ligament, spiral limbus, and stria vascularis were selected using freehand selections button. The “measure” function was used to obtain the mean gray scale value of the ROI, which was inversely correlated with the staining intensity. For confocal microscopy of immunofluorescence staining, the ROIs including stria basal cells, spiral ganglion cells, spiral ligament, and spiral limbus were extracted using photoshop CS6 (version 13.0, Adobe Systems Software Ireland Ltd, Dublin, Ireland) program and were imported into ImageJ program. The images were split into individual channel, and the green (corresponded to CD44 and TLR2) and red (corresponded to hyaluronic acid) channels were selected for further quantifications. The “Threshold” was adjusted using the “set” button in the “Image” menu, and “Limit to Threshold” option should be selected and “Direct to” should be defined to the corresponding channel in the “Analyze” menu. Then the gray scale value, which was inversely correlated with the staining intensity, was obtained using the “Measure” function in the same menu.
Statistical analyses were performed using the IBM ® SPSS ® Statistics Version 20 software package (SPSS Inc., Chicago, USA). A one-way ANOVA and Kruskal-Wallis test were used to compare ABR threshold shifts and signal intensities (grayscale) of staining for glycosaminoglycan and hyaluronic acid secretions, and TLR2 and CD44 staining between the LPN injected-ear and saline injected-ear groups in the different cochlear structures among various turns. Least significant difference (LSD) test was used as post hoc analysis. Higher numbers in the grayscale analysis correlate with lower signal intensities of the staining. p < 0.05 was accepted as statistically significant.
Abreviaturas
- ABR:
-
Auditory brainstem response
- AgNPs:
-
Silver nanoparticles
- dH2 O:
-
Deionized water
- GAB1:
-
Growth factor receptor-bound protein 2-associated-binding protein 1
- Gd-DOTA:
-
Gadolinium-tetra-azacyclo-dodecane-tetra-acetic acid (DOTAREM)
- Gd-MRI:
-
Gadolinium-enhanced magnetic resonance imaging
- JNK:
-
Jun amino-terminal kinases
- LPN + Gd-DOTA:
-
Gd-DOTA-containing LPNs
- LPNs:
-
Liposome nanocarriers
- ROI:
-
Region of interests
- ROS:
-
Especies reactivas de oxígeno
- TLR:
-
Toll-like receptor
Nanomateriales
- En sintonía con el corazón de un átomo de cobre
- Detección del magnetismo del núcleo de un solo átomo
- Imagen de carbono oceánico en escala atómica
- Después del crecimiento, llevar los sistemas de calibración atípicos al pliegue multinacional
- Después de 15 segundos, el cerebro olvida la información por completo
- Google encuentra su Glass después de dos años en la oscuridad
- El diseño de la capa de emisión para multiplicadores de electrones
- Efectos del pH microambiental de los liposomas sobre la estabilidad química del fármaco cargado
- Amazon-Whole Foods:dos años después del acuerdo
- Después del prototipo:la siguiente fase del proceso de diseño
- Vida útil de la bomba:después de la reconstrucción