


Nanoscopía basada en microscopía de fuerza atómica de células madre derivadas de tejido adiposo humano que se diferencian condrogenicamente:nanoestructura e integrina expresión β1
Resumen
Se sabe que la integrina β1 participa en la diferenciación, migración, proliferación, reparación de heridas, desarrollo de tejidos y organogénesis. Con el fin de analizar la probabilidad de unión entre el ligando de la integrina β1 y los receptores del grupo de diferenciación 29 (CD29), se utilizó microscopía de fuerza atómica (AFM) para detectar receptores acoplados a la integrina β1 nativa en la superficie de células madre derivadas de tejido adiposo humano (hADSc) . La probabilidad de unión de la interacción ligando-receptor de integrina β1 fue probada por puntas funcionalizadas de integrina β1 en hADSc durante la diferenciación condrogénica temprana a nivel de cultivo celular bidimensional. La morfología celular y la ultraestructura de hADSc se midieron mediante AFM, que demostró que las células con fusiones largas se convertían en células poligonales con relaciones de longitud / anchura disminuidas y una rugosidad aumentada durante la inducción condrogénica. La unión del ligando de la integrina β1 y los receptores CD29 se detectó mediante puntas funcionalizadas con β1 para hADSc vivo. Se registraron un total de 1200 curvas a los 0, 6 y 12 días de inducción condrogénica. Las fuerzas de rotura promedio fueron, respectivamente, 61,8 ± 22,2 pN, 60 ± 20,2 pN y 67,2 ± 22,0 pN. Los eventos de ruptura fueron 19,58 ± 1,74%, 28,03 ± 2,05% y 33,4 ± 1,89%, respectivamente, lo que demostró que la probabilidad de unión aumentaba entre el ligando de la integrina β1 y los receptores en la superficie de hADSc durante la inducción condrogénica. La integrina β1 y la ruta de señalización de β-catenina / SOX se correlacionaron durante la diferenciación condrogénica. Los resultados de esta investigación implican que AFM ofrece una visión cinética y visual de los cambios en la unión del ligando β1 de la integrina-receptor CD29 en hADSc durante la condrogénesis. Se demostró que los cambios en la morfología celular, la ultraestructura de la membrana y la probabilidad de unión ligando-receptor transmembrana son marcadores útiles para la evaluación del proceso de diferenciación condrogénica.
Antecedentes
La osteoartritis (OA) es una enfermedad común degenerativa de las articulaciones de los ancianos [1], con OA degenerativa caracterizada por la destrucción progresiva del cartílago articular. El cartílago está muy organizado sin vasos sanguíneos, nervios o tejido linfático [2]. La matriz extracelular (MEC) está compuesta principalmente por colágeno II y glucoproteína, y es muy importante para la homeostasis del cartílago. Dado que el cartílago es avascular, su capacidad de autorrenovación es limitada. Aunque los tratamientos de OA (tanto quirúrgicos como no quirúrgicos) pueden aliviar rápidamente los síntomas del paciente con OA, especialmente el dolor, no pueden restaurar la estructura y función normales del cartílago articular [3]. En el futuro, el tratamiento probablemente incluirá la ingeniería de tejidos con células madre y andamios para reparar defectos y cartílago articular degenerativo [4]. Las células madre mesenquimales son células estromales multipotentes que tienen potencial osteogénico, adipogénico, condrogénico y miogénico, dependiendo de las combinaciones de factores de crecimiento [5]. El análisis de la diferenciación de células madre mesenquimales ha demostrado que Wnt / β-catenina, diana de rapamicina en mamíferos (mTOR), fosfoinositido 3-quinasa (PI3K) y otras vías desempeñan funciones importantes en la diferenciación [6,7,8]. Sin embargo, el mecanismo subyacente por el cual se induce la diferenciación condrogénica sigue siendo difícil de alcanzar. Esto es particularmente cierto para el mecanismo por el cual las señales extracelulares activan las vías de señalización intracelular. Hemos encontrado que la integrina β1 experimenta cambios durante la diferenciación condrogénica. Por lo tanto, planteamos la hipótesis de que la integrina β1 puede desempeñar un papel importante en la diferenciación condrogénica de células madre derivadas de tejido adiposo humano (hADSc) debido a su participación en diversas vías de señalización de diferenciación tisular. En esta investigación, la atención se centró en la vía de señalización Wnt / β-catenina.
Numerosos estudios han demostrado que las interacciones entre las células y el entorno extracelular están reguladas por proteínas transmembrana, en particular, miembros de la familia de las integrinas [9]. Las integrinas están compuestas de glicoproteínas transmembrana heterodiméricas de cadenas α y β unidas no covalentemente [10]. En teoría, hay 64 integrinas conocidas de las cuales solo se han encontrado 24. Las integrinas juegan un papel vital en la adhesión célula-célula, la adhesión célula-ECM, la señalización celular y la organización del citoesqueleto de actina [11]. La ECM juega un papel importante en la homeostasis tisular y la ECM regula las integrinas. Las integrinas median muchos procesos fundamentales que incluyen la adhesión celular, la migración, la proliferación, la diferenciación, la muerte celular, la reparación de heridas, el desarrollo de tejidos y la organogénesis. Durante la diferenciación condrogénica de células madre mesenquimales, la expresión de la integrina β1 está conectada a la vía de señalización SOX y al colágeno II. El foco de esta investigación fue el dímero β1 de la integrina, ya que es el dímero β más prominente entre los heterodímeros del cartílago y se sabe que interactúa con muchos dímeros α diferentes [12]. El grupo de diferenciación 29 (CD29) es una subunidad de la integrina β1 asociada con receptores de antígenos muy tardíos, expresados en casi todas las células y tipos de tejidos.
Aquí, se utilizó microscopía de fuerza atómica (AFM) para ayudarnos a medir los cambios durante la diferenciación condrogénica de hADSc. Como un tipo de microscopía de sonda de barrido de muy alta resolución, el AFM ha brindado una nueva oportunidad para detectar la morfología y la membrana celular de células individuales en un líquido a nanoescala. Mientras tanto, se utilizó el sistema de espectroscopía de fuerza de una sola molécula (SMFS) combinado por microscopía de fuerza atómica (AFM) para medir la unión ligando-receptor en las células vivas. El sistema de SMFS era más sensible a los cambios de receptores en la membrana celular y se visualizaron las imágenes de la fuerza de unión. En este trabajo, la unión del receptor-ligando de la integrina β1 fue probada por las puntas de AFM funcionalizadas con la integrina β1. Aplicando AFM, se encontró que la diferenciación condrogénica cambia la forma de las células hADSc y aumenta la rugosidad celular. Esta aplicación proporcionó un método para evaluar la diferenciación condrogénica mediante la medición directa de las interacciones ligando-receptor de la integrina β1 y la alteración de la ultraestructura de la superficie celular, mejorando la investigación de la superficie celular y el cribado de forma visualizada. La diferenciación condrogénica cambia la composición y estructura de la membrana, así como las interacciones citoesqueléticas intracelulares. Estos cambios en la morfología celular, la ultraestructura y la unión ligando-receptor transmembrana sirven como marcadores útiles para la evaluación de los mecanismos de diferenciación condrogénica.
Métodos
Cultivo celular y reactivos
Para esta investigación, se aislaron células de tres pacientes quirúrgicos (edad media de 20 años) como se describió anteriormente [13]. Se obtuvieron los consentimientos informados de todos los pacientes. La aprobación ética para este estudio se obtuvo del Primer Hospital Afiliado de la Universidad de Jinan (formulario de suplemento). Las células se mantuvieron en medio basal, que incluía medio de Eagle modificado de Dulbecco bajo en glucosa (DMEM, Life Technologies, CA, EE. UU.) Suplementado con suero bovino fetal inactivado por calor al 10% (FBS, Life Technologies, CA, EE. UU.), 100 unidades / ml de penicilina (Life Technologies, CA, EE. UU.), 100 μg / ml de estreptomicina (Life Technologies, CA, EE. UU.), 0,11 mg / ml de piruvato de sodio (Life Technologies, CA, EE. UU.) y L-glutamina (Life Technologies, CA, EE.UU). Las células se mantuvieron a 37 ° C en una incubadora humidificada que contenía CO 2 al 5%. con medio cambiado cada 3 días.
Diferenciación in vitro
Para la inducción condrogénica, se sembraron hADSc de cuarto a octavo pasaje a una densidad celular alta (2 × 105/10 ml) y se cultivaron en medio condrogénico que contenía DMEM / F12 suplementado con FBS al 1%, insulina-transferrina-selenio al 1% ( ITS) + suplemento (Cyagen, Guangzhou, China), 10 ng / ml de factor de crecimiento transformante beta1 (TGF-β1) (Peprotech, Rocky Hill, Nueva Jersey, EE. UU.), 100 ng / ml de factores de crecimiento similares a la insulina-1 ( IGF-1) (Peprotech, Peprotech, Rocky Hill, Nueva Jersey, EE. UU.), Dexametasona 10-7 M (Sigma, St. Louis, MO, EE. UU.) Y 50 μg / ml de ácido ascórbico (Sigma, St. Louis, MO , EE.UU). El medio se cambió cada 2 días con TGF-β1 e IGF-1 recién añadidos. La condrogénesis se evaluó mediante tinción con azul alcián y azul de toluidina.
Para inducir la diferenciación osteogénica y adipogénica, las células del cuarto al octavo pasaje se trataron con el medio osteogénico y adipogénico durante 2 semanas, respectivamente. El medio osteogénico consistió en DMEM suplementado con dexametasona 10-7 M (Sigma, St. Louis, MO, EE. UU.), 50 μg / ml de ácido ascórbico (Sigma, St. Louis, MO, EE. UU.) Y 10 mmol / l de β-glicerol fosfato (Sigma, St. Louis, MO, EE. UU.). La osteogénesis se evaluó mediante tinción con rojo de alizarina.
El medio adipogénico consistía en DMEM suplementado con 0,5 mmol / l de 3-isobutil-1-metilxantina (IBMX) (Sigma, St. Louis, MO, EE. UU.), 1 μmol / l de hidrocortisona (Sigma, St. Louis, MO, EE. UU.), 0,1 mmol / l de indometacina (Sigma, St. Louis, MO, EE. UU.). La diferenciación adipogénica se evaluó mediante tinción Oil Red O.
Identificación de antígenos de superficie hADSc por citometría de flujo
Las hADSC se digirieron con tripsina y luego se enjuagaron dos veces con DMEM, antes de resuspenderlas a una densidad celular de 2 × 10 7 células / ml. La suspensión celular (50 μl; 1 × 10 6 células) se añadió a tubos de epoxi epóxido de 1,5 ml y luego se incubó con anticuerpos anti-CD34, anti-CD44, anti-CD45, anti-CD73, anti-CD90, anti-CD106, anti-HLA-DR y anti-CD105 para 20 min a 37 ° C en la oscuridad. Los anti-CD34, anti-CD44 y anti-CD45 se obtuvieron de CST (Beverly, MA, EE. UU.); otros anticuerpos se obtuvieron de Abcam (Cambridge, MA, EE. UU.). Luego, la suspensión celular se centrifugó a × 500 g durante 5 min, seguido de la eliminación del sobrenadante y la resuspensión de las células en 200 l de tampón de tinción. Todos los pasos se repitieron dos veces antes del análisis por citometría de flujo.
Análisis de inmunotransferencia (IB)
Las células se recolectaron para inmunotransferencia como se describió anteriormente [14]. Los anticuerpos primarios utilizados fueron anti-β-catenina (ab32572), antiintegrina β1 (ab30394) y anti-colágeno II (ab34712), obtenidos de Abcam (Cambridge, MA, EE. UU.). Anti-β-actina (8H10D10, 1:2000), anti-GSK-3β (27C10, 1:1000) y anti-SOX (92G2, 1:1000) se obtuvieron de Cell Signaling Technology (CST, Beverly, MA, EE.UU). Los anticuerpos secundarios conjugados con HRP (1:1000-1:3000) se compraron en CST.
Inmunofluorescencia
Para la diferenciación condrogénica, las células se trataron durante 0, 6 y 12 días, se digirieron y se cultivaron en el vidrio en placas de 24 pocillos (Costar353047, Corning, Nueva York, EE. UU.) Durante 24 h. Las células se lavaron dos veces con solución tampón de fosfato (PBS) enfriada con hielo, se fijaron con paraformaldehído al 4% durante 15 min a temperatura ambiente. Después del bloqueo, las células se incubaron con el anticuerpo primario reactivo con integrina β1 durante 1 h, seguido de incubación durante 1 h en la oscuridad con IgG anti-ratón (H + L) marcado con Alexa Fluor 488 (CST # 4408, MA, EE. UU. ), 4 ', 6-diamidino-2-fenilindol (DAPI, Sigma, MO, EE. UU.). Para la tinción con faloidina, después del bloqueo, las células se permeabilizaron con Triton X-100 al 0.2% durante 30 min, luego las células se incubaron con DAPI y faloidina-Alexa Flour 573 (Life Technologies, CA, EE. UU.) Durante 1 h. Después de lavar tres veces, se evaluó la localización subcelular de la integrina β1 y el cambio de actina filamentosa (F-actina) durante la diferenciación del cartílago con un microscopio confocal de barrido láser (ZEISS, LSM 700, Oberkochen, Alemania).
Preparación de consejos de AFM
Las puntas de Si3N4 (DNP-10, Bruker Corp) con una constante de resorte (0,06 N / m) fueron modificadas químicamente por el anticuerpo anti-CD29 como sigue [15]. Las puntas se limpiaron con acetona, luz ultravioleta y solución de piraña (H 2 SO 4 :H 2 O 2 =3:1, v / v ) para diferentes tiempos (5 min, 30 min y 10 min). Después de enjuagar a fondo con agua purificada, las puntas se formaron mediante incubación con una solución de 3-APTES al 1% (Sigma, St. Louis, MO, EE. UU.) En etanol durante 30 min. Las puntas se lavaron con agua ultrapura tres veces y se trataron con una solución de glutaraldehído al 2,5% (Sigma, St. Louis, MO, EE. UU.) Durante 1 h. El glutaraldehído superfluo se lavó tres veces con agua. Finalmente, las puntas se insertaron en una solución anti-integrina β1 (1 mg / ml) y se incubaron durante la noche a 4 ° C. Las sondas modificadas se lavaron con PBS antes de los experimentos.
Medidas de AFM
Se utilizó AFM (Bioscope Catalyst, Bruker, EE. UU.) Para investigar la morfología de hADSc y los cambios ultraestructurales durante la diferenciación condrogénica. La constante de fuerza exacta de las puntas de AFM se midió en PBS. Para evaluar la morfología y la ultraestructura, las células se lavaron con PBS varias veces. Luego, se añadió una solución de paraformaldehído al 4% en un 2 de 3,5 cm. plato de cultivo durante 15 min. Después de lavar las células con PBS, las células se almacenaron en PBS a 4ºC hasta su uso. La constante elástica de las puntas osciló entre 4,2 y 5,8 N / m en el modo de contacto. Se tomaron imágenes morfológicas y ultraestructurales de hADSc en PBS a temperatura ambiente por AFM. Las imágenes de ultraestructura que rodean los núcleos de hADSc se obtuvieron en modo de contacto. Se utilizó un software de análisis de nanoscopio para evaluar la ultraestructura de la superficie celular para más de 15 imágenes diferentes de 10 × 10 μm para al menos 15 células diferentes en grupos (día 0, 6, 12). Se analizó la fuerza de unión entre las puntas de AFM modificadas con integrina β1 y los receptores CD29 de hADSc viva durante diferentes períodos condrogénicos (0, 6 y 12 días). La fuerza de unión se midió en el modo de aproximación-retracción del sistema AFM (Bioscope Catalyst, Bruker, EE. UU.). Para estudiar los eventos de separación de células vivas de integrina β1, se utilizaron las puntas modificadas con anticuerpos de integrina β1 a velocidades de aproximación-retracción de 500 nm / s. La constante de fuerza de las puntas funcionalizadas fue de 0,058 ± 0,006 N / m. La fuerza umbral sobre las células fue de 800 pN. El anticuerpo anti-integrina β1 (100 µg / ml) se añadió a las células durante 30 minutos antes de los experimentos de medición de fuerza. El bloqueo de integrina β1 y las sondas desnudas también se utilizaron como controles para detectar la fuerza de ruptura inespecífica entre las puntas y las células modificadas con anticuerpo de integrina β1. Para la cuantificación de la probabilidad de unión del ligando-receptor de la integrina β1, se midieron las curvas de fuerza de interacción específica mediante sondas funcionalizadas con el anticuerpo de la integrina β1. Se midieron más de 400 curvas de fuerza en un solo experimento con resultados resumidos de al menos tres experimentos independientes. Por lo tanto, se adquirieron aproximadamente 1200 curvas originales de fuerza-distancia en cada experimento de comparación de 30 a 40 células diferentes utilizando el software de análisis Nanoscope del instrumento. Al promediar los valores de fuerza para al menos tres experimentos independientes, se determinó el efecto de la inducción condrogénica sobre la fuerza de interacción entre el ligando de la integrina β1 y los receptores CD29 en la superficie celular.
Transcripción inversa y PCR en tiempo real
Se utilizaron kits de purificación de ARN TRIzol® Plus (Life Technologies, CA, EE. UU.) Y se realizó la transcripción inversa de 1 μg de ARN a ADNc utilizando un equipo de transcripción inversa de ADNc de alta capacidad (Invitrogen) de acuerdo con el protocolo del fabricante con modificaciones menores. La integrina β1 y GAPDH se cuantificaron usando qRT-PCR con cebadores específicos de genes:5′-TGGAGGAAATGGTGTTTGC-3 ′ (integrina β1-sentido) y 5′-CGTTGCTGGCTTCACAAGTA-3 ′ (integrina β1 antisentido); 5′-CTGACTTCAACAGCGACACC-3 ′ (GAPDH-sentido) y 5′-CCCTGTTGCTGTAGCCAAAT-3 ′ (GAPDH-antisentido). Para la PCR en tiempo real, la PCR en tiempo real del paso uno (Applied Biosystems) se realizó utilizando Fast SYBR @ GREEN Master Mix (Life Technologies, CA, EE. UU.). La expresión del gen diana se normalizó a GAPDH como patrón interno y se calculó usando el método comparativo 2-ΔΔCT. Cada ensayo se realizó por triplicado.
Análisis estadístico
Todos los experimentos se realizaron al menos tres veces, con los datos expresados como media ± desviación estándar (DE). t realizó la comparación entre dos grupos prueba. Las diferencias significativas entre las medias de los grupos se determinaron mediante un análisis ANOVA de una vía, seguido de la prueba T2 de Bonferroni y Tamhane (no se asumieron variaciones iguales). Valores de p <0,05 se consideraron estadísticamente significativos.
Resultados y discusión
Evaluación de hADSc
Las células madre mesenquimales son células estromales multipotentes que tienen potencial osteogénico, adipogénico, condrogénico y miogénico. Hay dos medios principales para identificar hADSc, los marcadores de CD de la superficie celular y la capacidad de diferenciar [16]. Como se muestra en el archivo adicional 1:Figura S1 y el archivo adicional 2:Figura S2, las células derivadas fueron hADSc. Luego, la proliferación celular del paso 3 hADSc se determinó mediante el ensayo MTT (archivo adicional 3:Figura S3).
Morfología inducida y cambios en la ultraestructura de la superficie durante la condrogénesis de hADSc
El AFM siempre se utiliza para detectar la morfología celular y la ultraestructura a nanoescala [17]. La forma de una célula se relaciona con su función celular especializada y con la organización de los tejidos. En algunas investigaciones sobre el cáncer, el AFM se puede utilizar como una técnica de alta formación de imágenes para analizar los cambios morfológicos para la evaluación de los efectos de los fármacos. Además, la forma de las células madre mesenquimales cambia durante la inducción condrogénica [18]. Si bien los cambios en la forma de las células parecen ser necesarios para la diferenciación, se sabe poco acerca de si la morfología celular afecta las etapas de desarrollo más tempranas de la diferenciación de las células madre mesenquimales. Por lo tanto, la AFM evaluó los cambios en la morfología y la ultraestructura de la membrana durante la condrogénesis de hADSc, ya que estos cambios son importantes [19] y pueden influir directamente en la función de las células [20]. Se investigaron la morfología de la superficie y la estructura ultrafina de hADSc durante la diferenciación condrogénica durante diferentes períodos de tiempo (Fig. 1 y Fig. 2). La morfología y la ultraestructura de la superficie fueron obviamente diferentes en cada grupo de comparación. El día 0, las células tenían una forma de huso alargado con una superficie relativamente lisa. La arquitectura de la membrana celular fue homogénea. Después de la inducción condrogénica, en los días 6 y 12, se observaron cambios significativos en la morfología celular. La mayoría de las células se contrajeron gradualmente a una forma poligonal (Fig. 1a) con una disminución en la proporción promedio de longitud / anchura de las células durante la diferenciación condrogénica (Fig. 1b). Numerosos estudios muestran que los cambios de la morfología celular son compatibles con el citoesqueleto de las células [21]. También encontramos cambios en el citoesqueleto durante la diferenciación condrogénica, lo que se explica en los últimos resultados.
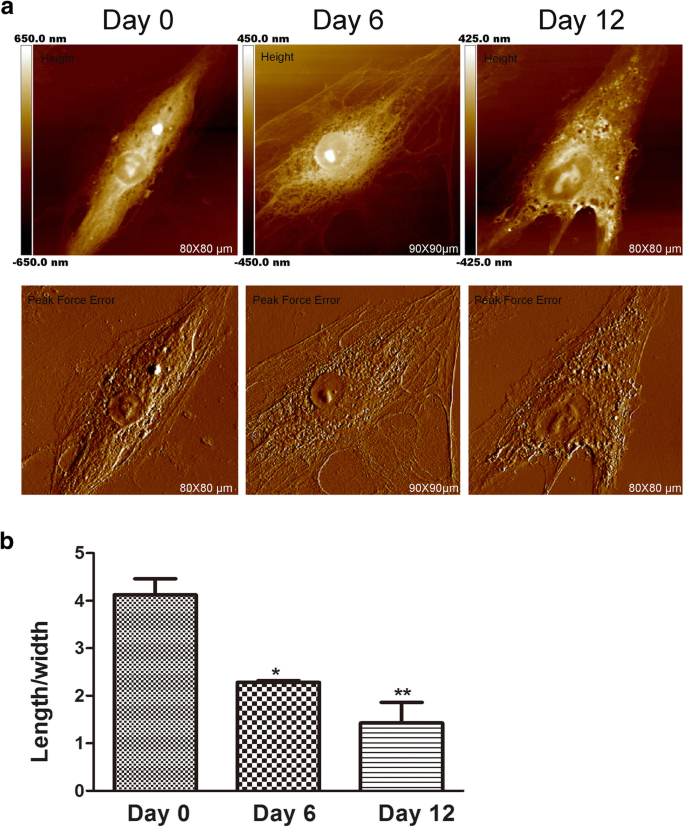
Características de la morfología de hADSc durante la condrogénesis. un Se obtuvieron imágenes morfológicas de hADSc completo a los 0, 6 y 12 días de diferenciación condrogénica. Las imágenes se analizaron mediante un modelo de imagen de error de fuerza máxima y altura mediante un nanoscopio. b Se midió la relación longitud / ancho promedio de las células después del tratamiento de diferenciación condrogénica a los 0, 6 y 12 días. * p <0.05, ** p <0.01
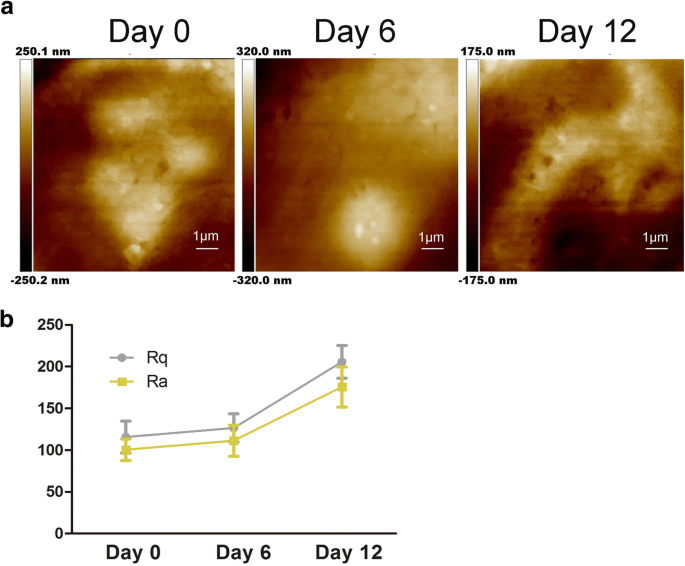
Características de la ultraestructura de la membrana hADSc durante la diferenciación condrogénica. un Los cambios en la ultraestructura de la membrana celular se evaluaron después de la diferenciación condrogénica durante 0, 6 y 12 días. b Los parámetros de rugosidad de la superficie Ra y Rq de las células se midieron durante la inducción condrogénica de hADSc durante 0, 6 y 12 días
Como se muestra en la Fig. 2a, la ultraestructura de la membrana celular también cambió; las partículas se agrandaron y fueron heterogéneas. Estudios anteriores demostraron que Ra y Rq fueron los creadores del valor de rugosidad para evaluar el cambio en las membranas celulares tratadas de manera diferente [22]. Rq trata sobre la rugosidad de la raíz cuadrada media, \ (\ mathrm {Rq} =\ sqrt {\ frac {\ sum_ {t-1} ^ N {\ left (Zn- \ overline {Z} \ right)} ^ 2 } {N-1}} \); \ (\ mathrm {Rq} =\ sqrt {\ frac {\ sum _ {\ mathrm {t} -1} ^ {\ mathrm {N}} {\ left (\ mathrm {Zn} - \ overline {\ mathrm {Z }} \ right)} ^ 2} {\ mathrm {N} -1}}; \) Ra tiene una rugosidad promedio, \ (\ mathrm {Ra} =\ frac {1} {N} {\ sum} _ { t-1} ^ N1 \ mid Zi- \ overline {Z} \ mid \). Para obtener la rugosidad, el tamaño de escaneo es de 10 μm × 10 μm. Como se muestra en la Fig. 2b, tanto el Ra como el Rq de dos áreas diferentes aumentaron durante la condrogénesis de hADSc. Los valores de Ra y Rq de las células en el día 0 eran bajos, lo que indica una superficie lisa (Fig. 2b). Los valores de Ra y Rq aumentaron simultáneamente con la diferenciación condrogénica, mostrando una mayor heterogeneidad y más rugosa en las superficies celulares (Fig. 2a). Según los cambios observados, la diferenciación condrogénica dio como resultado la morfología celular y cambios en la relación altura / ancho de la célula (Fig. 1a, b). Hay estudios que muestran que la ECM podría regular la adhesión celular mediante la regulación de las integrinas [11]. Por lo tanto, el aumento de los valores de rugosidad sugirió cambios en la ECM y la ultraestructura de la membrana celular durante la condrogénesis. Estos datos demuestran que la diferenciación condrogénica afecta la morfología celular, la ECM y la estructura de la membrana celular.
Cambios citoesqueléticos durante la inducción condrogénica de hADSc
Durante la diferenciación de las células madre, la morfología celular y los cambios estructurales de la membrana están relacionados con el citoesqueleto de la célula, después del desarrollo de características celulares específicas del linaje [21]. Como se muestra en la Fig. 3a, las señales de fluorescencia roja y azul indican respectivamente F-actina y DAPI. El citoesqueleto celular cambió mucho durante la inducción condrogénica en la Fig. 3a. Por un lado, los microfilamentos del citoesqueleto fueron a lo largo del eje celular largo en el grupo del día 0, mientras que los microfilamentos del citoesqueleto se extendieron en una matriz radial cuando las hADSc se trataron con diferenciación condrogénica durante 12 días. Por otro lado, la distribución de los microfilamentos celulares fue homogénea en el grupo del día 0, pero los microfilamentos se distribuyeron principalmente en la periferia de hADSc tratados con diferenciación condrogénica durante 12 días.
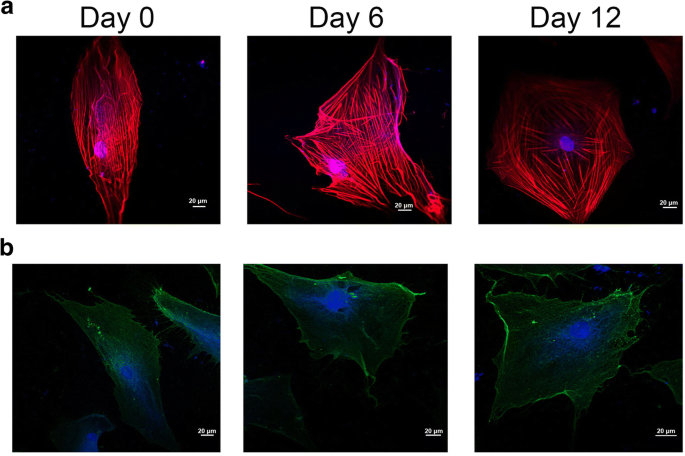
Organización del citoesqueleto y ubicación de la integrina β1 en hADSc que se diferencian condrogenicamente. un Se detectaron cambios en el citoesqueleto durante la condrogénesis de hADSc mediante microscopía confocal. b La ubicación de la integrina β1 se midió durante la diferenciación condrogénica mediante microscopía confocal. El citoesqueleto y el núcleo se tiñeron con F-actina y DAPI, respectivamente. Las señales de fluorescencia roja y azul indican respectivamente F-actina y DAPI
La diferenciación condrogénica cambió la probabilidad de unión de la integrina β1 a los receptores en hADSc
El AFM también es una herramienta útil para el estudio de la fuerza de unión entre los ligandos y sus receptores, lo que aclara la transducción de la señal del receptor de la membrana en la superficie celular [23]. Mediante AFM, los cambios entre la integrina β1 y sus receptores se miden de forma visual, simple y específica. La interacción del receptor-ligando de la integrina β1 en las células vivas es una forma de explorar el proceso de unión en la membrana celular. El procedimiento para la funcionalización de la punta de AFM es el acoplamiento de la integrina β1 a las puntas de AFM mediante la unión de APTES y glutaraldehído. Estas puntas se usaron para la detección de la unión de la integrina β1 a los receptores CD29 en las superficies celulares (Fig. 4a). Se utilizó espectroscopia de fuerza de molécula única (SMFS) para evaluar la distribución de la fuerza de separación de células vivas antiintegrina β1 dentro de regiones localizadas de hADSc vivas individuales (Fig. 4b). Las curvas de fuerza representativas se muestran en la Fig. 4c, d, que representan una curva de una sola molécula (Fig. 4c) y dos pares de curvas de pico de ruptura (Fig. 4d). Se realizaron experimentos de bloqueo y experimentos con puntas de AFM desnudas para verificar la especificidad de las curvas de fuerza obtenidas. Las puntas de AFM desnudas no detectaron un pico de fuerza específico (Fig. 4e). Los experimentos de AFM desnudos mostraron que la probabilidad de unión inespecífica de la interacción ligando-receptor de la integrina β1 en la superficie de hADSc era inferior al 1%. Para los experimentos de bloqueo, el anticuerpo anti-integrina β1 se incubó con células durante 30 min y luego se registraron las curvas de fuerza usando puntas funcionalizadas con integrina β1. El anticuerpo de bloqueo redujo las curvas de fuerza en un 90% (Fig. 4f). No hubo diferencia en la probabilidad de unión del ligando-receptor de la integrina β1 en las superficies celulares entre los tres grupos después del tratamiento con anticuerpos anti-integrina β1 (Fig. 4g). Estos resultados demuestran que las puntas de AFM modificadas con anticuerpos fueron muy útiles para detectar la fuerza y que las puntas de AFM funcionalizadas con integrina β1 eran específicas.
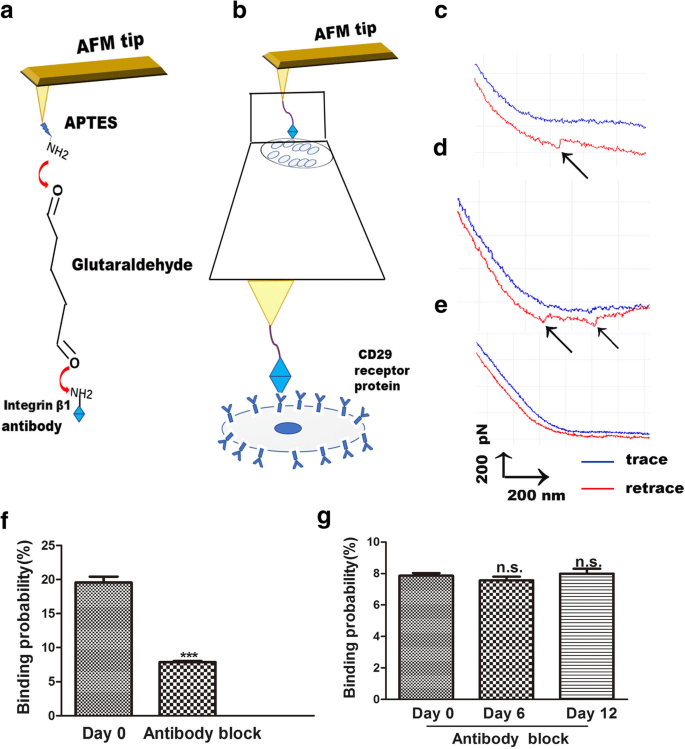
Medidas de fuerza de AFM con punta de AFM funcionalizada con integrina β1 en hADSc vivo. un Representación esquemática de la estrategia utilizada para la inmovilización de la integrina β1 en una punta de AFM. b Representación esquemática de la fuerza de una sola molécula medida entre las puntas de AFM funcionalizadas con integrina β1 y hADSc vivo. c, d Curvas de fuerza representativas obtenidas con puntas de AFM modificadas con integrina β1 en hADSc y e después de que el sistema se bloqueó con la solución de anticuerpo monoclonal de integrina β1. f La probabilidad de unión de las puntas funcionalizadas con integrina β1 en hADSc antes y después del bloqueo por el anticuerpo de integrina β1 en el día 0. g La probabilidad de unión de puntas funcionalizadas con CD29 en hADSc después del bloqueo por el anticuerpo de la integrina β1 a los 0, 6 y 12 días. *** p <0.001, n.s .. sin diferencia significativa
La fuerza de unión (fuerza de ruptura) es la fuerza de interacción entre los ligandos y sus receptores [24]. Los cambios en la morfología y la ultraestructura de la superficie de las membranas plasmáticas están relacionados con muchos procesos de la biología celular, como la diferenciación, la apoptosis y la migración celular. Durante la diferenciación, se cree que los cambios en el citoesqueleto están relacionados con los cambios de la integrina, especialmente la integrina β1. La integrina β1 (CD29) es muy importante en la adhesión celular a la ECM y en la adhesión célula-célula. También puede interactuar con proteínas intracelulares, estimulando moléculas de señalización relacionadas con el citoesqueleto de actina [25]. En este estudio, se observaron cambios en la morfología celular y citoesquelética durante la condrogénesis de hADSc mediante microscopía de barrido láser confocal (CLSM) y AFM. Durante la diferenciación condrogénica, los cambios en el citoesqueleto, la morfología y la ultraestructura de la superficie pueden ser un indicador nuevo y confiable del estado celular. La integrina β1, el receptor CD29, se distribuye sobre la superficie celular según se juzga por inmunofluorescencia (Fig. 3b). La fuerza de unión y la estabilidad de los complejos ligando-receptor de la integrina β1 durante la condrogénesis de hADSc se evaluaron a los 0, 6 y 12 días de diferenciación. Se registraron un total de 1200 curvas por día, con fuerzas de ruptura promedio de 61,8 ± 22,2 pN, 60 ± 20,2 pN y 67,2 ± 22,0 pN, respectivamente (fig. 5a-c). La distribución de la magnitud de la fuerza se analizó como fuerza media + SD (figura 5d). No hubo diferencia significativa en la media de fuerza entre los días 0 y 6. Hubo una diferencia en la media de fuerza entre los días 0 y 12. La magnitud de la fuerza de unión aumentó el día 12. Mientras tanto, eventos de ruptura a los 0, 6 y 12 días fueron, respectivamente, 19,58 ± 1,74%, 28,03 ± 2,05% y 33,4 ± 1,89% (Fig. 5e). La mayor probabilidad de unión también indicó que la integrina β1 (CD29) jugó un papel importante en la diferenciación condrogénica y puede proporcionar la información para la diferenciación condrogénica, a través de vías de señalización. Por tanto, el aumento de los nanodominios β1 de la integrina durante la diferenciación condrogénica puede afectar fundamentalmente la fuerza de unión del ligando-receptor CD29 en hADSc vivo. Los cambios en la morfología y la ultraestructura de la superficie de las membranas plasmáticas acompañaron a los cambios en la estructura, conformación, fuerza de unión y estabilidad de la proteína de la integrina β1 en las células. En resumen, la integrina β1 juega un papel necesario en la diferenciación condrogénica de hADSc.
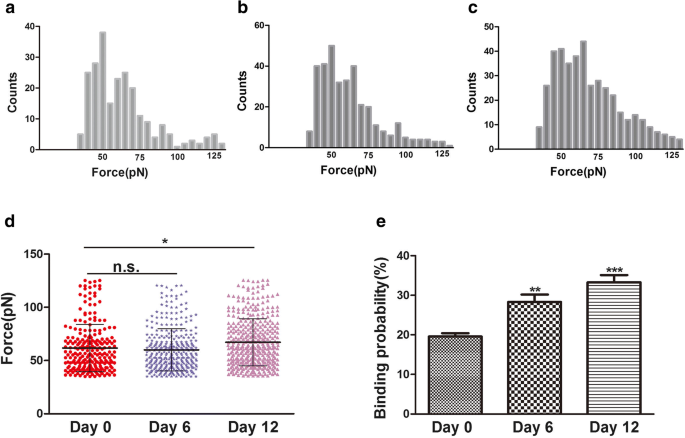
Fuerza de unión y probabilidad de unión medidas en la superficie de hADSc vivo mediante puntas de AFM funcionalizadas con integrina β1. a – c Histogramas de la fuerza de unión del anticuerpo-receptor de la integrina β1 obtenidos durante la diferenciación condrogénica de hADSc durante 0, 6 y 12 días. d Las fuerzas de unión para los receptores de integrina β1 se obtuvieron a los 0, 6 y 12 días de diferenciación condrogénica de hADSc. e La probabilidad de unión del receptor de integrina β1 se detectó durante la diferenciación condrogénica de hADSc durante 0, 6 y 12 días. * p <0.05, ** p <0.01, *** p <0,001, n.s. sin diferencia significativa
Regulación al alza de la integrina β1 durante la diferenciación condrogénica de hADSc
Numerosos estudios han demostrado que los miembros de la familia de las integrinas juegan un papel importante en la diferenciación celular. Además, las integrinas pueden regular la interacción entre el entorno extracelular y las células, controlando las vías de transducción de señales a través de proteínas conectadas [26]. Estudios anteriores han demostrado que la probabilidad de unión puede verse afectada por la densidad y conformación de la proteína transmembrana (receptores) en la superficie celular [27]. La conformación de integrina puede ser un casco cerrado, que tiene baja afinidad por el ligando, o un casco abierto, que tiene una alta afinidad por el ligando [28, 29]. La expresión de la integrina β1 se reguló positivamente tanto a nivel transcripcional como traduccional con una expresión aumentada de colágeno II, característica de los condrocitos (Fig. 6a, b). Como tal, la expresión de integrina β1 regulada positivamente fue consistente con una mayor probabilidad de unión sin tener en cuenta la conformación.

The role of integrin β1 and β-catenin/SOX pathway in regulating hADSc chondrogenic differentiation. un Protein integrin β1 was up-regulated during chondrogenesis of hADSc as assessed by western blotting. Cartilage differentiation up-regulated collagen II expression at different days. b The mRNA of integrin β1 was up-regulated during chondrogenic differentiation of hADSc. c Measurement of proteins associated with the β-catenin/SOX pathway during chondrogenic differentiation of hADSc for 0, 6, and 12 days. * p <0.05, ** p < 0.01
The Role of Integrin β1 in Chondrogenic Differentiation Regulated by the β-catenin/SOX Signaling Pathway
Previous studies have shown Wnt/β-catenin, PI3K, and mTOR signaling pathways to be related to integrin β1 [30,31,32]. Each is important in mesenchymal stem cell differentiation. Likewise, studies have demonstrated SOX and collagen II to be regulated by integrin β1 during chondrogenesis of hADSc. SOX is a hallmark component of the Wnt/β-catenin signaling pathway. Hence, we hypothesized that chondrogenic differentiation was regulated by the β-catenin/SOX pathway via integrin β1. SOX, GSK-3β, β-catenin, and integrin β1 were all increased during chondrogenesis of hADSc (Fig. 6c), with integrin β1 inducing cell signaling. These data demonstrate chondrogenic differentiation to be regulated by the β-catenin/SOX pathway via integrin β1.
Prospective and Limitations
In this work, changes in cellular morphology, the structure of the membrane, and the binding probability of integrin β1 ligand–receptors were demonstrated to be useful image markers to evaluate the chondrogenic differentiation process. This is a new method for evaluation of morphology, membrane ultrastructure, and changes in transmembrane proteins during chondrogenic differentiation. There are limitations to this study. Although increased binding probability was related to the high expression of integrin β1, the conformation of integrin β1 during chondrogenesis was not investigated. Further work is necessary to determine the conformation of integrin β1 during chondrogenic differentiation. Integrin β1 was demonstrated to participate in the β-catenin/SOX signaling pathway during chondrogenesis of hADSc. However, the relationship between integrin β1 and β-catenin/SOX signaling pathway is still not fully established. Further work is necessary to identify the exact role of integrin β1 in this pathway.
Conclusiones
In the present work, a novel method (AFM) was employed to evaluate chondrogenic induction in hADSc. Cell surface ultrastructural changes were assessed by AFM imaging. AFM was used to investigate the binding force and binding probability between integrin β1 ligand and its receptors on the surface of hADSc by integrin β1-functionalized AFM tips. Based on AFM data, during chondrogenesis, cell morphology was changed from an elongated spindle shape to a polygonal shape with increased cell roughness. By use of integrin β1-functionalized AFM tips, the binding probability and force magnitude of integrin β1 ligand–receptor on the surface of hADSc were found to increase during chondrogenic induction. By immunoblot, integrin β1 was demonstrated to participate in the β-catenin/SOX signaling pathway, which regulated the chondrogenesis of hADSc. Taken together, these results and the established methodology contribute to a better understanding of cell morphology and roughness. Further, the data provide thermodynamic and kinetic insight into the integrin β1 ligand-binding process, at the single-molecule level. This AFM method will be useful for investigation of signaling pathways in living hADSc during chondrogenesis. Changes in the cellular nanostructure, as well as structure of the membrane, and the binding probability of transmembrane proteins are useful markers to evaluate chondrogenic differentiation mechanisms. This AFM method can be used to understand the mechanism of mesenchymal stem cell differentiation in tissue engineering and will be useful for an enhanced understanding of mesenchymal stem cell chondrogenic differentiation.
Abreviaturas
- AFM:
-
Microscopía de fuerza atómica
- CD:
-
Cluster of differentiation
- DAPI:
-
4′,6-diamidino-2-phenylindole
- DMEM:
-
Dulbecco’s Modified Eagle’s Medium
- ECM:
-
Extracellular matrix
- F-actina:
-
Actina filamentosa
- FBS:
-
Suero fetal bovino
- hADSc:
-
Human adipose-deprived stem cells
- IB:
-
Immunoblotting analysis
- IBMX:
-
3-isobutyl-1-methylxanthine
- IGF-1:
-
Insulin-like growth factors-1
- ITS:
-
Insulin transferrin selenium
- mTOR:
-
Mammalian target of rapamycin
- OA:
-
Osteoarthritis
- PBS:
-
Solución tampón de fosfato
- PI3K:
-
Phosphoinositide 3-kinase
- SD:
-
Desviación estándar
- SMFS:
-
Single-molecule force spectroscopy
- TGF-β1:
-
Transforming growth factor-beta1
Nanomateriales
- Demostración de un biosensor flexible basado en grafeno para la detección rápida y sensible de células de cáncer de ovario
- Tecnologías avanzadas de deposición de capas atómicas para micro-LED y VCSEL
- Nanopartículas de fósforo negro promueven la diferenciación osteogénica de EMSC a través de la expresión TG2 regulada hacia arriba
- Presentación de la estructura atómica y electrónica de las nanofibras de carbono de copa apilada
- Análisis de actina y organización de la adhesión focal en células U2OS en nanoestructuras poliméricas
- Las nanotecnologías desactivan las células madre cancerosas
- Comparación de electrodos metálicos estampados tipo nanoagujero y tipo nanopilar incorporados en células solares orgánicas
- Síntesis de nanocristales de ZnO y aplicación en células solares de polímero invertido
- Estudio de la fuerza de adhesión y la transición vítrea de películas delgadas de poliestireno mediante microscopía de fuerza atómica
- Una sonda fluorescente reanudable BHN-Fe3O4 @ SiO2 Nanoestructura híbrida para Fe3 + y su aplicación en bioimagen
- Dielectroforesis de ondas de frecuencia modulada de vesículas y células:cambios de sentido periódicos en la frecuencia de cruce