


Impacto de las nanopartículas de oro en el metabolismo de la testosterona en microsomas hepáticos humanos
Resumen
Los complejos corona de nanopartículas de oro (AuNP) -proteína pueden alterar el metabolismo de la testosterona (TST) mediada por el citocromo P450 (CYP) al alterar sus propiedades fisicoquímicas. Investigamos el impacto del tamaño de NP, la química de la superficie y la corona de proteínas en el metabolismo de la TST en microsomas hepáticos humanos agrupados (pHLM) empleando AuNP de 40 y 80 nm funcionalizado con polietilenimina ramificada (BPEI), ácido lipoico (LA) y polietilenglicol (PEG). ) así como la corona de proteínas plasmáticas humanas (PC). También se caracterizó la variación individual en el metabolismo de TST mediado por AuNP entre HLM de un solo donante que contenían diferentes niveles de actividades de CYP. Se produjeron efectos inhibidores de AuNP de 40 nm y, en menor grado, de AuNP de 80 nm para la producción de un total de cinco metabolitos hidroxilados de TST en pHLM, pero la PC los alivió. Mientras tanto, AuNP desnudo aumentó la producción de androstenediona. La variación interindividual en el metabolismo de la TST se produjo dentro de la HLM de un solo donante. En la mayoría de los casos, 40 y 80 nm desnudo y PC AuNP suprimieron esencialmente el metabolismo de la TST a una concentración no inhibidora, pero PC PEG-AuNP aumentó la androstenediona. Estos estudios contribuyen a una mejor comprensión del papel de AuNP como disruptor de TST al alterar el metabolismo de TST y podrían utilizarse para detectar otros NP como disruptores endocrinos potenciales.
Introducción
Las nanopartículas de oro (AuNP) se han utilizado ampliamente en la administración de fármacos, el diagnóstico médico y la terapia del cáncer, así como en productos de consumo, es decir, cosméticos, envases de alimentos, debido a sus propiedades ópticas y físicas únicas [1, 2, 3]. Tras la exposición a una mezcla de proteínas, las NP se asocian con proteínas y forman una corona de proteínas, que altera la química de la superficie, las conformaciones de una proteína adsorbida y las respuestas biológicas posteriores, es decir, toxicidad de NP, absorción de NP celular, actividad catalítica del citocromo P450 (CYP ) enzimas hacia fármacos [4, 5, 6, 7]. Los estudios in vitro con células epiteliales primarias y líneas de células cancerosas sugieren que AuNP era tóxico para los hepatocitos humanos, la línea de células de hepatoma C3A y los espermatozoides [6, 7, 8]. Pero la formación de corona de proteínas alrededor de NP atenuó intensamente o potenció la toxicidad de AuNP de una manera dependiente de la química de la superficie [6, 7]. La proteína corona interfirió la captación celular de AuNP en hepatocitos humanos, células del túbulo proximal renal, células HepG2, línea celular C3A, independientemente de sus tamaños y cargas superficiales [6, 7, 9,10,11,12].
Las enzimas CYP hepáticas están involucradas principalmente en la síntesis y / o el metabolismo de compuestos endógenos y exógenos, pero una amplia gama de agentes, es decir, fármacos, pesticidas o NP, afectan de forma inversa la síntesis, el metabolismo y / o la desintoxicación de las hormonas esteroides dando lugar a efectos farmacológicos. y la función fisiológica [13,14,15,16,17]. La testosterona (TST) es un andrógeno importante y sustrato específico de CYP3A4 (una conversión principal a 6β-OH TST) de una manera regio y estereo-selectiva [18]. Durante la fase I del metabolismo, la TST también es hidroxilada a 2β-OH TST por CYP3A4 y desalquilada a androstenediona (AD) por CYP2D6 [17, 19]. Estudios in vitro con hepatocitos humanos, línea celular C3A, microsomas hepáticos humanos (HLM) y enzimas CYP recombinantes sugirieron que AuNP desnudo y recubierto de corona de proteína modulaba una amplia gama de enzimas CYP que incluían CYP1A2, 2C9, 2C19, 2D6, 2E1, y 3A4 [6, 7, 20, 21]. Otras NP metálicas, AgNP desnudas, también suprimieron la producción mediada por CYP3A4 de 6β-OH TST en HLM [22]. AuNP funcionalizado con polietilenimina ramificada (BPEI) y ácido lipoico (LA) disminuyó la actividad de CYP3A4 en la línea celular C3A, pero la corona de proteína plasmática humana (PC) la atenuó [7]. Por el contrario, los BPEI-AuNP desnudos (sin PC) y PC fueron inhibidores de CYP2C9 y 3A4 en hepatocitos humanos, independientemente del tamaño de NP [6].
Los estudios in vivo informaron que un tamaño pequeño de AuNP (4 y 13 nm) se acumuló principalmente en el hígado y el bazo en ratones macho BALB / c e indujo la expresión de los genes hepáticos Cyp1a1 y 2b [23]. Otros NP metálicos, NP de óxido de zinc, inhibieron la actividad hepática CYP1A2, 2C11 y 3A2 en ratas macho Sprague Dawley con un aumento de los cambios patológicos en el hígado [24].
Hasta la fecha, se sabe poco cómo las AuNP asocian el metabolismo de la TST mediado por CYP (hidroxilación y desalquilación de la TST) en ausencia y / o presencia de una corona proteica biológicamente relevante. Los objetivos de este estudio son investigar el impacto de la PC en las propiedades fisicoquímicas de la AuNP BPEI catiónica de 40 y 80 nm, la AuNP LA aniónica y la AuNP de polietilenglicol neutro (PEG). El impacto de AuNP en el metabolismo de TST mediado por CYP con y sin PC se caracterizará utilizando pHLM. También se estudiará la variación individual en el metabolismo de la TST en el HLM de un solo donante que contenía varios grados de enzimas CYP.
Métodos / Experimental
Productos químicos
El 2,3,4- 13 C 3 testosterona (CAS # 327048-83-9) y 6β-hidroxitestosterona (6β-OH TST, CAS # 62-99-7) se obtuvieron de MilliporeSigma (St. Louis, MO). Testosterona (TST, CAS # 58-22-0), 2α-hidroxitestosterona (2α-OH TST, CAS # 004075-14-3), 2β-hidroxitestosterona (2β-OH TST, CAS # 10390-14-4), 6α -hidroxitestosterona (6α-OH TST, CAS # 2944-87-8), 11β-hidroxitestosterona (11β-OH TST, CAS # 1816-85-9), 15β-hidroxitestosterona (15β-OH TST, CAS # 39605-73- 7), 16α-hidroxitestosterona (16α-OH TST CAS # 63-01-4), 16β-hidroxitestosterona (16β-OH TST, CAS # 17528-90-4), 11-cetotestosterona (CAS # 564-35-2) , androstenediona (AD, CAS # 63-05-8), 4-hydroxy androstenediona (CAS # 566-48-3) y 11β-hydroxy androstenediona (CAS # 382-44-5) se adquirieron de Steraloids (Newport, RI ). El acetonitrilo y el ácido fórmico de grado LC-MS se obtuvieron de Fisher Scientific (Fair Lawn, NJ), mientras que el agua ultrapura se produjo internamente mediante el sistema Synergy® UV-R de Merck KGaA (Darmstadt, Alemania). Si no se especifica, todos los demás reactivos se compraron a MilliporeSigma (St. Louis, MO).
Microsomas de hígado humano
Se obtuvieron microsomas de hígado humano agrupados (pHLM) (200 donantes, 100 machos y 100 hembras) y microsomas de hígado de un solo donante de Corning Inc. (Charlotte, NC). Los pHLM son agrupados por el proveedor, pero no un grupo de un solo HLM de donante. Las características y la actividad enzimática del citocromo P450 (CYP) seleccionado del HLM de donante único utilizado en este estudio se presentan en el archivo adicional 1:Tabla S1.
Síntesis de nanopartículas de oro
Se adquirieron Biopure ™ AuNP esférico de 40 y 80 nm funcionalizado con polietilenimina ramificada catiónica (BPEI), ácido lipoico aniónico (LA) y polietilenglicol neutro (PEG) de nanoComposix (San Diego, CA). Los materiales del núcleo se sintetizaron mediante la reducción del hidrato de tetracloroaurato (III) de hidrógeno (HAuCl 4 3H 2 O) en solución acuosa de carbonato de potasio y se sometió al proceso de envejecimiento y filtración de flujo tangencial (TFF). La superficie de AuNP se funcionalizó con LA o PEG añadiendo ácido dihidrolipoico (0,2:1, w / w ) o PEG terminado en tiol-metoxi (Laysan Bio Inc., Arab, AL) (0,5:1, w / w ), respectivamente, con lavado TFF y filtración estéril. Las superficies funcionalizadas con BPEI se sintetizaron mediante la química de EDC / NHS uniendo el ácido carboxílico de LA a aminas de BPEI. El BPEI no unido se eliminó con lavado de TFF y una centrifugación posterior.
Preparación de corona de proteínas plasmáticas humanas
Plasma humano combinado (HP, n =5) se obtuvo de Biological Specialty Corp. (Colmar, PA). La AuNP de 40 y 80 nm se incubó con plasma humano a un volumen de plasma fisiológico en volumen de sangre total, 55% ( v / v ) en una incubadora orbital con agitación / rotación a 37 ° C a 250 rpm durante 1 h. Al final de la incubación, la corona de proteínas plasmáticas humanas (PC) que rodea al NP se recogió mediante centrifugación a 20.000 × g a 20 ° C durante 20 min seguido de tres lavados con solución salina tamponada con fosfato (PBS). Las proteínas no unidas y unidas débilmente se descartaron mediante centrifugación. Las PC AuNP resultantes se dispersaron en PBS y se utilizaron para caracterizar las propiedades fisicoquímicas y su interacción con TST.
Caracterización física de AuNP
El tamaño de partícula y las propiedades de la superficie se midieron mediante dispersión dinámica de luz (DLS) y microscopía electrónica de transmisión (TEM). Diámetros hidrodinámicos (D H ), y el potencial zeta de 40 y 80 nm desnudos (sin PC) BPEI-, LA- y PEG-AuNP en agua desionizada (DI) y PC AuNP en PBS se midieron con el Zetasizer Nano-Zs (Malvern Instruments, Worcestershire, Reino Unido) a 0 ha 25 ° C. La D H , índice de polidispersidad (PDI) y potencial zeta también se obtuvieron para AuNP desnudo y PC en un tampón de incubación microsomal (pH 7,4) a 0 min y 45 min a 37 ° C. Las muestras se midieron cinco veces con 11 sub-corridas de 10 s cada una. TEM caracterizó la morfología de AuNP desnudo y PC. Todos los AuNP se colocaron en rejillas de cobre recubiertas de formvar y se visualizaron en un Tecnai G2 Spirit BioTWIN con un detector Oxford (FEI Company, Hillsboro, OR) a un voltaje de aceleración de 120 kV. La suite de microscopía GATAN (GATAN Inc., Pleasanton, CA) midió los diámetros de AuNP. Se midió un espectro de absorción óptica con el lector de microplacas multimodo Spectra Max i3 (Molecular Devices, Sunnyvale, CA).
Metabolismo in vitro de la testosterona en ausencia y presencia de AuNP desnudo y PC
Se realizaron estudios preliminares para determinar el tiempo de incubación y las concentraciones de proteína microsomal para proporcionar una tasa metabólica lineal para TST (una concentración final de 10 μM). La producción de metabolitos de TST fue lineal de 1.3 a 9.3 mg de proteína microsomal mL −1 por hasta 60 min. Los ensayos metabólicos se realizaron como se describe en su totalidad [25]. Brevemente, se trató pHLM en un tampón de incubación microsomal con TST 10 μM y posteriormente, se agregaron AuNP desnudas (sin PC) de 40 y 80 nm a 0, 7, 32, 63, 143, 250, 400 y 571 μg mL −1 ; para PC AuNP pHLM 0, 7, 32, 63 y 143 μg mL −1 . Un tampón de incubación microsomal contenía tampón fosfato 100 mM, MgCl 2 3,3 mM y EDTA 1 mM (pH 7,4). La reacción metabólica se inició con y sin un sistema de regeneración de NADPH que contenía NADP 0,25 mM, glucosa-6-fosfato 2,5 mM y 2 U mL −1 glucosa-6-fosfato deshidrogenasa. Después de 45 min de incubación a 37 ° C, la reacción se detuvo agregando 4% ( v / v ) solución acuosa de ácido fosfórico (1:1, v / v ). Después de una centrifugación a 3500 rpm durante 20 min, se recogió un sobrenadante de muestra y se almacenó a -20 ° C hasta su uso posterior. Además, el HLM de un solo donante en un tampón de incubación microsomal se trató con TST 10 μM seguido de incubación con 63 μg mL −1 de todos los desnudos y PC AuNP durante 45 min a 37 ° C. Al final de la incubación, la muestra se procesó y se almacenó a - 20 ° C como se indicó anteriormente.
Preparación de estándares y muestras
Soluciones madre estándar primarias de TST, sus metabolitos y 13 C 3 Se prepararon TST marcadas como patrón interno (ISTD) en metanol a una concentración de 1 mM y se almacenaron a -20 ° C hasta su uso posterior. Las concentraciones para las soluciones estándar de trabajo de TST y sus metabolitos fueron 0.01, 0.05, 0.1, 0.5, 1, 5, 10, 50, 100 y 200 μM con diluciones seriadas de la solución madre primaria. Para los calibradores estándar, se añadió una alícuota de 50 μL de cada solución estándar de trabajo a 450 μL de tampón de reacción, lo que dio como resultado diluciones 1:10, mientras que también se preparó una solución ISTD de 0,1 μM utilizando una solución acuosa de ácido fosfórico al 4%. Se prepararon muestras de control de calidad (QC) a concentraciones de 0.01, 0.05 y 0.1 μM.
Después de descongelar, las muestras se sometieron a centrifugación a 3500 rpm durante 20 min a temperatura ambiente. El sobrenadante se enriqueció con 50 μL de 0,1 μM de ISTD y se sometió a una placa de elución de 96 pocillos Oasis PRIME HLB y una placa de recolección en el procesador Waters de presión positiva-96 a 80 psi durante 1-2 min. (Waters Corp., Milford, MA). Después de lavar con 300 μL de agua con metanol al 5% y eluir con 50 μL de mezcla de acetonitrilo / metanol (90/10, v / v ), el eluyente resultante se diluyó en 50 μL de agua (un volumen final de 100 μL) y se sometió a cromatografía líquida-espectrometría de masas (LC-MS / MS).
Cromatografía líquida-Espectrometría de masas
Todas las muestras se separaron en una columna Waters UPLC HSS T3 (2,1 x 50 mm, 1,8 μm) con el sistema de cromatografía líquida Waters Acquity Ultra Performance con detector de triple cuadrupolo (UPLC TQD) (Waters Corp., Milford, MA). Las fases móviles A y B eran ácido fórmico al 0,1% en agua y ácido fórmico al 0,1% en metanol, respectivamente. Se utilizó un método de LC en gradiente a una velocidad de flujo de 600 μL min −1 durante 0–8,4 min. El gradiente fue de 0 a 1 min (30% B), 1 a 3 min (a 50% B), 3 a 3,5 min (50% B), 3,5 a 7 min (a 80% B), 7 a 7,01 min ( al 98% B), 7,01–7,5 min (98% B) y 7,51–8,4 min (30% B). Las condiciones de la EM se describen brevemente a continuación. La fuente de ionización se hizo funcionar en electropulverización positiva (ESI + ) modo con voltaje capilar 4000 V; para la temperatura de la fuente 150 ° C; y para temperaturas de desolvatación 450 ° C. Los caudales de gas de desolvatación (N 2 ), gas de cono (N 2 ) y el gas de colisión (argón) fueron 900 L h −1 , 100 L h −1 y 0,1 ml min −1 , respectivamente. El tipo de exploración fue la monitorización de reacciones múltiples (MRM) y el tiempo de ejecución de MS fue de 8,4 min. Las transiciones de MRM utilizadas para el análisis se resumieron en el archivo adicional 1:Tabla S2 y Fig. 2. El volumen de inyección fue de 2 μL y la columna se mantuvo a 50 ° C durante todo el análisis. Todos los métodos de cuantificación se basaron en una curva de calibración de siete puntos en rangos de concentración de 0,001 a 20 μM. El límite de detección (LOD) y el límite de cuantificación (LOQ) de 0,001 μM y 0,005 μM se establecieron para TST y metabolitos diana.
Análisis estadístico
Los efectos de los dispersantes sobre D H y el PDI de desnudo y PC en AuNP se evaluaron utilizando la t de Student prueba con una distribución de dos colas. Concentración inhibitoria media máxima (IC 50 ) y la mitad de la concentración máxima de activación (EC 50 ) de AuNP hacia la producción de metabolitos de TST dependientes de CYP en pHLM se determinaron ajustando una ecuación de Hill con pendiente variable a los datos observados usando GraphPad Prism®. Se realizó un análisis de varianza unidireccional (ANOVA) utilizando GraphPad Prism® para evaluar los efectos del tratamiento con AuNP sobre el metabolismo de la TST en HLM de un solo donante. Cuando los efectos fueron significativos, se realizó una comparación múltiple con la prueba de diferencia significativa honesta (HSD) de Tukey al 5% de nivel de significancia. Coeficiente de correlación de Pearson ( r ) entre la actividad CYP de HLM de un solo donante y la producción de metabolitos de TST dependientes de CYP se determinó utilizando GraphPad Prism® versión 6.07 (La Jolla, CA).
Resultados y discusión
Caracterización fisicoquímica de la corona de proteína plasmática humana y desnuda AuNP
El impacto de la corona de proteínas plasmáticas humanas (PC) en el tamaño de NP, la carga superficial y la morfología, así como una propiedad espectral, se ha caracterizado utilizando espectroscopía DLS, TEM y UV-Vis (Fig. 1). Las imágenes TEM demostraron que todas las AuNP desnudas (sin PC) y PC AuNP, excepto las BPEI-AuNP PC de 40 y 80 nm, eran monodispersas con una distribución de tamaño constante y rangos de espectro UV-Vis únicos (520-557 nm) (Fig. 1a, b) . TEM también encontró PC distinta alrededor de AuNP en PBS. La agregación de la polietilenimina ramificada (BPEI) -AuNP revestida con PC de 40 y 80 nm en PBS a 0 min a 25 ° C correlacionó múltiples picos en la distribución de tamaño y corrimientos al rojo de los espectros de absorción con respecto a la BPEI-AuNP desnuda (Fig. 1b). El diámetro hidrodinámico (D H ) Los valores de PC BPEI-AuNP de 40 y 80 nm disueltos en PBS a 0 min a 25 ° C y en tampón de incubación microsomal a 0 y 45 min a 37 ° C no fueron determinados por DLS, junto con múltiples picos en la distribución de tamaño. La D H Los valores de BPEI- y LA-AuNP desnudos de 40 nm y PC PEG-AuNP y el BPEI-AuNP desnudo de 80 nm en tampón de incubación microsomal aumentaron sustancialmente hasta 45 min a 37 ° C, mientras que su valor disminuyó para el PC LA de 80 nm -AuNP (Tabla 1). El índice de polidispersidad (PDI) de 40 nm desnudo y PC PEG-AuNP y 80 nm PC PEG-AuNP aumentó a los 45 min a 37 ° C. Además, los valores de potencial zeta (z) del BPEI-AuNP desnudo de 40 y 80 nm y del PEG-AuNP desnudo de 40 nm disminuyeron sustancialmente con el tiempo. Un estudio anterior informó que AuNP (7 y 70 nm) asociado con proteínas microsomales del hígado humano alteraron el máximo característico de absorbancia en el rango UV-visible [21]. Estos resultados fueron apoyados por los estudios recientes en nuestro laboratorio que la PC y la corona de albúmina sérica humana alteraron el tamaño de la NP, el desplazamiento al rojo de la absorbancia máxima y la morfología independientemente del medio de disolución y el tiempo de incubación [6, 7, 10, 26]. Sobre la base de los cambios en las propiedades fisicoquímicas de NP mediadas por PC y la función enzimática de las actividades de CYP [6, 7], se investigó el efecto potencial de AuNP de 40 y 80 nm en el metabolismo de la TST microsomal hepática humana mediada por CYP en presencia de un factor más biológico PC relevante.

Micrografías electrónicas de transmisión de ( a ) el AuNP desnudo de 40 y 80 nm en agua desionizada y b el AuNP de PC de 40 y 80 nm en PBS a 0 ha 25 ° C, el espectro de absorción de UV (recuadro superior) y la distribución de dispersión de luz dinámica (recuadro inferior). Las flechas indican la formación de PC. BPEI polietilenimina ramificada, LA ácido lipoico, PEG polietilenglicol, ND no determinado, PC corona de proteína plasmática humana, desnuda sin PC
Metabolismo de testosterona mediado por AuNP en microsomas hepáticos humanos agrupados
Se examinaron un total de 11 metabolitos de TST y se encontraron seis metabolitos en microsomas hepáticos humanos agrupados (pHLM) a TST 10 μM. El análisis LC-MS / MS dirigido de TST y los seis metabolitos seleccionados se muestra en la Fig. 2. La lista de los metabolitos seleccionados incluía cinco metabolitos de TST hidroxilados (2β-OH TST, 6β-OH TST, 15β-OH TST, 16α -OH TST, 16β-OH TST) y un metabolito desalquilado (androstenediona, AD). Esto se correlaciona con los estudios anteriores que utilizaron hepatocitos humanos y HLM de que la TST se hidroxilaba principalmente en 6β-OH TST y, en menor grado, 2β-OH TST, 15β-OH TST, 16α-OH TST y 16β-OH TST, así como un metabolito desalquilado, AD [17, 19, 27].
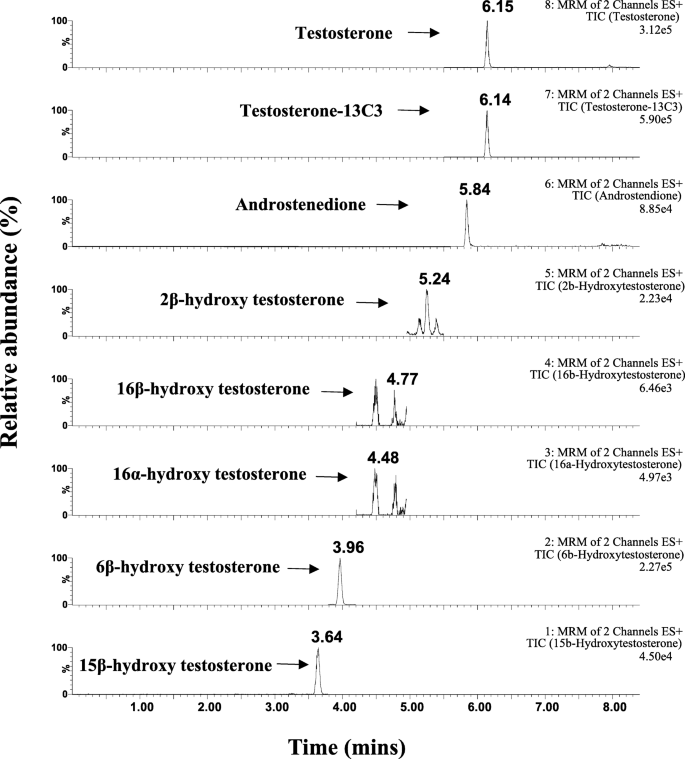
El cromatograma de iones extraído (XIC) para testosterona (TST), 13 TST marcada con C3, androstenediona, 2β-hidroxitestosterona (2β-OH TST), 16α-hidroxitestosterona (16α-OH TST), 16β-hidroxitestosterona (16β-OH TST), 6β-hidroxitestosterona (6β-OH TST) y 15β -hidroxitestosterona (15β-OH TST) producida en HLM combinado a la concentración final de testosterona 10 μM en presencia de NADPH durante 45 min a 37 ° C. HLM microsomas de hígado humano, NADPH un fosfato de dinucleótido de nicotinamida y adenina reducido
El resultado de la coincubación de TST con AuNP desnudo de 40 y 80 nm en pHLM se muestra en las Figs. 3 y 4. Todas las AuNP desnudas de 40 y 80 nm alteraron la producción de 2β-OH TST, 6β-OH TST y 15β-OH TST en pHLM con un grado variable de inhibición (Fig. 3a-f). Concentración inhibitoria media máxima (IC 50 ) Los valores de BPEI-AuNP de 40 nm para la producción de 6β-OH TST fueron 416 μg mL −1 ; para 80 nm BPEI-AuNP 438 μg mL −1 ; y para PEG-AuNP de 80 nm 387 μg mL −1 (Figura 3c, d). Para la producción de 15β-OH TST, IC 50 Los valores de BPEI-AuNP de 40 nm fueron 1113 μg mL −1 ; para BPEI-AuNP de 80 nm 415 μg mL −1 ; y para PEG-AuNP de 80 nm 480 μg mL −1 (Figura 3e, f). Estos resultados fueron respaldados por estudios in vitro con líneas de células cancerosas humanas y tejidos hepáticos en los que un NP metálico, AgNP y NP de silicio poroso impidieron la producción de 6β-OH TST en células Caco2 de adenocarcinoma colorrectal epitelial humano, células HepG2 de carcinoma hepatocelular y microsomas hepáticos humanos [22, 28]. El AuNP desnudo de 40 y 80 nm no inhibió la producción de 16α-OH TST y 16β-OH TST, excepto por el BPEI-AuNP de 40 nm que suprimió la producción de 16β-OH TST en la concentración más alta (517 μg mL - 1 ) (Fig. 4a – d). El BPEI-AuNP de 40 y 80 nm aumentó la producción de androstenediona (AD) en la concentración más alta con las tasas metabólicas correspondientes de 4,3 y 2,1 pmol mg de proteína −1 min −1 , respectivamente en comparación con el control (0,7 pmol mg de proteína −1 min −1 ) (Figura 4e, f). Pero el LA-AuNP de 80 nm inhibió la producción de AD con IC 50 valor de 233 μg mL −1 . Estos resultados indicaron que el AuNP desnudo mediaba la producción del metabolito TST seleccionado en el revestimiento de la superficie y de maneras dependientes del tamaño. Además, las PC AuNP de 40 y 80 nm no inhibieron la producción de seis metabolitos seleccionados de TST en pHLM hasta 143 μg mL −1 , independientemente de los revestimientos superficiales (Figs. 5 y 6). Especialmente, la PC alivió la inhibición mediada por BPEI-AuNP desnuda de 40 nm de la producción de 6β-OH TST y 15β-OH TST a concentraciones más altas (32 μg mL −1 a 143 μg mL −1 ) (Figuras 3c, e y 5c, e). Estos resultados mostraron que el BPEI-, LA- y PEG-AuNP desnudos de 40 y 80 nm disminuyeron la hidroxilación de TST (2β-OH TST, 6β-OH TST y 15β-OH TST) de una manera dependiente de la dosis (Fig.7 ). Además, el BPEI-AuNP desnudo de 40 y 80 nm aumentó la producción de AD, pero el primero disminuyó la 16β-OH TST. Un estudio in vitro informó que la producción de 6β-OH TST mediada principalmente por CYP3A4 fue inhibida por nanotubos de carbono de pared simple (SWCNT) de una manera dependiente de la dosis, pero la corona de albúmina de suero bovino la alivió [17, 29]. Nuestro laboratorio informó recientemente que el BPEI-AuNP desnudo y PC de 40 y 80 nm sirvió como inhibidor de CYP1A2, 2C9 y 3A4 a nivel celular y transcripcional [6, 7]. Un estudio in vivo informó que la PEG-AuNP (4 y 13 nm) se acumuló principalmente en el hígado de ratones machos BALB / c y se alteraron los niveles transcripcionales de los genes hepáticos Cyp1a1 y 2b [23]. Ratones ICR macho con i.v. inyección de PEG-NH 2 -AuNP exhibió NP aumento del nivel de TST en plasma sin la morfología y fertilidad de los espermatozoides [30]. Otro NP metálico, dióxido de titanio (TiO 2 ) se acumuló en los testículos de ratones machos CD1 y disminuyó la expresión de cyp1b1 y 2e1 [31]. El estudio epidemiológico informó que un hombre adulto en la clínica de infertilidad de Massachusetts mostró niveles bajos de TST en plasma comparados con un alto nivel de 3,5,6-tricloro-2-piridinol (TCP) derivado de una alta exposición a clorpirifos (SFC), un conocido disruptor endocrino y un inhibidor del metabolismo de la TST mediado por CYP [32]. Un estudio anterior informó que los insecticidas disruptores endocrinos conocidos, CFS, CFS oxon, fonofos, forato, dietiltoluamida (DEET) y permetrina inhibieron sustancialmente y / o activaron la producción de metabolitos de TST hidroxilados y / o desalquilados, es decir, 2β-OH TST, 6β-OH TST, 15β-OH TST y AD y 4-hidroxi AD en hígado humano [17]. Dicho esto, es razonable postular que AuNP puede ser un disruptor endocrino potencial al mediar una potencia inhibidora y / o activadora contra el metabolismo de la TST mediado por CYP.
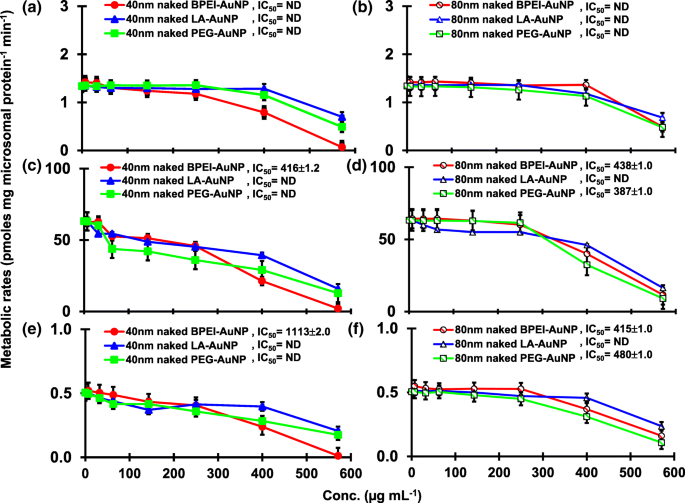
Efecto inhibidor de AuNP desnudo (sin PC) sobre la producción de 2β-OH TST ( a , b ), 6β-OH TST ( c , d ) y 15β-OH TST ( e , f ) en pHLM. Los datos representan la media ± S.D. ( n =3). IC 50 , la mitad de la concentración inhibitoria máxima; pHLM , microsomas de hígado humano agrupados; ND , no determinado ajustando una ecuación de Hill con pendiente variable a los datos observados usando GraphPad Prism®; PC , proteína plasmática humana; BPEI polietilenimina ramificada; LA , ácido lipoico; PEG, polietilenglicol; Conc, concentración; 2β-OH TST, 2β -hidroxitestosterona; 6β-OH TST, 6β -hidroxitestosterona; 15β-OH TST, 15β -hidroxitestosterona
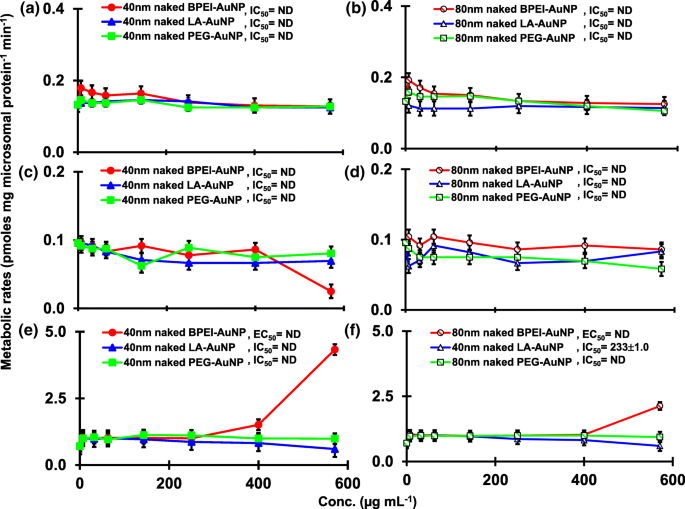
Un efecto inhibidor y estimulante de AuNP desnudo (sin PC) sobre la producción de 16β-OH TST ( a , b ), 16β-OH TST ( c , d ) y AD ( e , f ) en pHLM. Los datos representan la media ± S.D. ( n =3). IC 50 , la mitad de la concentración inhibitoria máxima; EC 50 , la mitad de la concentración máxima de activación; pHLM , microsomas de hígado humano agrupados; ND , no determinado ajustando una ecuación de Hill con pendiente variable a los datos observados usando GraphPad Prism®; PC , proteína plasmática humana; BPEI polietilenimina ramificada; LA , ácido lipoico; PEG , polietilenglicol; Conc, concentración; 16α-OH TST, 16α -hidroxitestosterona; 16β-OH TST, 16β -hidroxitestosterona; AD , androstenediona.
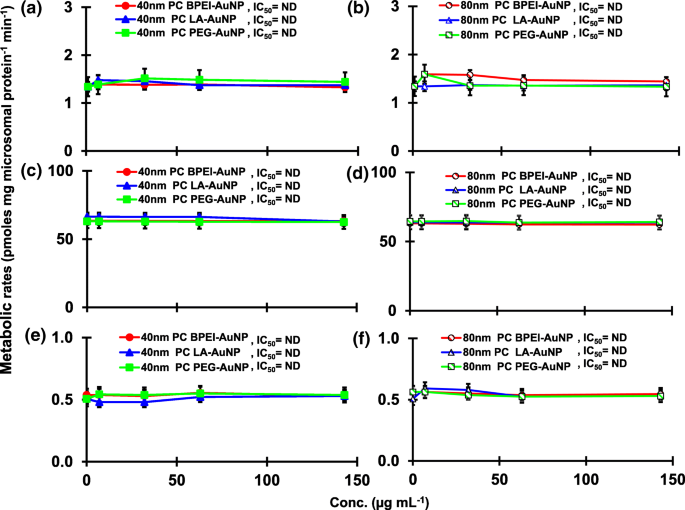
Efectos de la PC AuNP sobre la producción de 2β-OH TST ( a , b ), 6β-OH TST ( c , d ) y 15β-OH TST ( e , f ) en pHLM. Los datos representan la media ± S.D. ( n =3). IC 50 , la mitad de la concentración inhibitoria máxima; pHLM , microsomas de hígado humano agrupados; ND , no determinado ajustando una ecuación de Hill con pendiente variable a los datos observados usando GraphPad Prism®; PC , corona de proteínas plasmáticas humanas; BPEI polietilenimina ramificada; LA , ácido lipoico; PEG , polietilenglicol; Conc, concentración; 2β-OH TST, 2β -hidroxitestosterona; 6β-OH TST, 6β -hidroxitestosterona; 15β-OH TST, 15β-hidroxitestosterona.
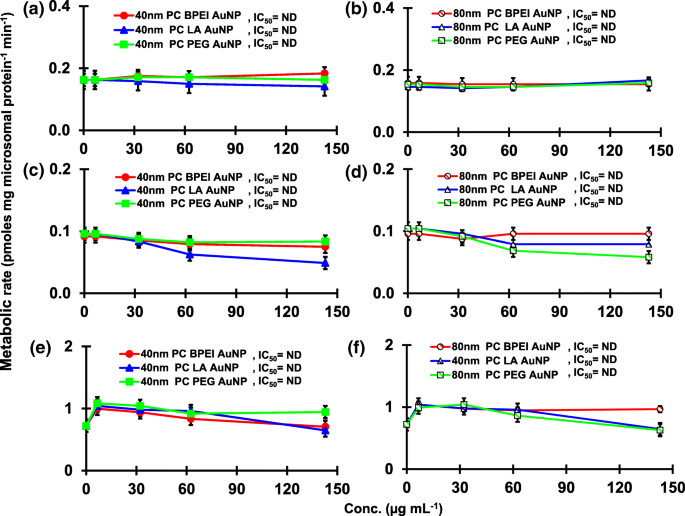
Efectos de la PC AuNP sobre la producción de 16α-OH TST ( a , b ), 16β-OH TST ( c , d ) y AD ( e , f ) en pHLM. Los datos representan la media ± S.D. ( n =3). IC 50 , la mitad de la concentración inhibitoria máxima; pHLM , microsomas de hígado humano agrupados; ND , no determinado ajustando una ecuación de Hill con pendiente variable a los datos observados usando GraphPad Prism®; PC , corona de proteínas plasmáticas humanas; BPEI polietilenimina ramificada; LA , ácido lipoico; PEG , polietilenglicol; Conc, concentración; 16α-OH TST, 16α -hidroxitestosterona; 16β-OH TST, 16β -hidroxitestosterona; AD , androstenediona.
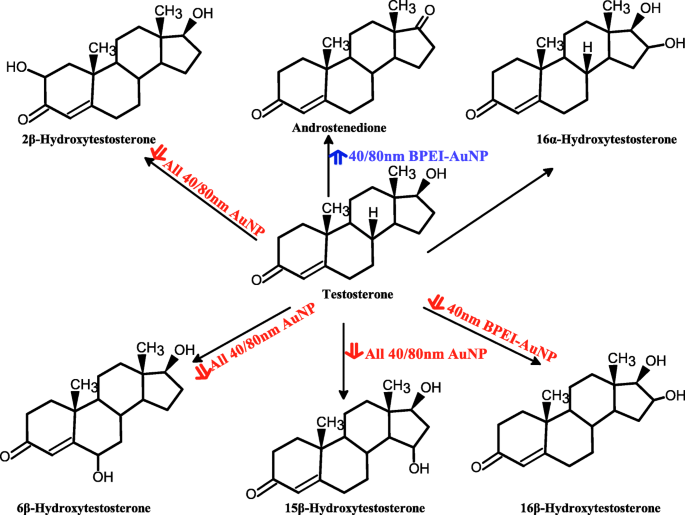
Esquema propuesto de metabolismo de la testosterona en microsomas hepáticos humanos agrupados y producción de metabolitos mediada por AuNP. AuNP nanopartículas de oro, BPEI polietilenimina ramificada, LA ácido lipoico, PEG polietilenglicol, PC corona de proteínas plasmáticas humanas. Flecha roja, efecto inhibidor; flecha azul, efecto estimulante
Metabolismo de la TST en microsomas hepáticos humanos de un solo donante y su modulación por AuNP
La producción de seis metabolitos seleccionados se caracterizó con HLM de un solo donante aislado de donantes con grado variable de actividad CYP a la concentración no inhibidora basada en pHLM de 40 y 80 nm desnudo y PC AuNP (10 μg mL −1 ). Se observó una variación individual en el metabolismo de la TST entre tres diferentes HLM de un solo donante (archivo adicional 1:Figura S1). La relación entre la actividad catalítica de cada enzima CYP y la producción de metabolitos derivados de TST se caracterizó en tres diferentes HLM de un solo donante que contenían una actividad catalítica CYP baja, media y alta (Archivo adicional 1:Tabla S1). La producción de 6β-OH TST se correlacionó positivamente con la actividad de CYP2C19 ( r =0,99 y p =0.01) y CYP3A4 ( r =0,99 y p =0.03) dentro de los individuos (Archivo adicional 1:Figura S2). La producción de AD se correlacionó negativamente con CYP4A11 ( r =- 0,98 y p =0.04) (Archivo adicional 1:Figura S3). Estos resultados fueron consistentes con un estudio previo que informó que CYP3A4 y CYP2D6 desempeñaron un papel clave en la producción de un metabolito principal de la TST, la 6β-OH TST y la EA, respectivamente [17]. Este estudio también sugirió que una variación individual en el metabolismo de TST dependiente de CYP depende de los genotipos de las enzimas CYP y sus actividades fenotípicas. Un estudio anterior informó que los polimorfismos y fenotipos de CYP son las características clave en la función de CYP y dan lugar a fenotipos farmacogenéticos categóricos como metabolizadores lentos, intermedios, extensos y ultrarrápidos que contribuyen a la susceptibilidad individual a reacciones adversas a los medicamentos y / o eficacia del medicamento y sugerencia de dosis. ese pobre metabolizador de la enzima CYP, es decir, CYP3A4, puede ser susceptible al metabolismo de la TST por la exposición al inhibidor de CYP, AuNP [33].
Como se muestra en las Figs. 8 y 9, la coincubación de TST con AuNP a una concentración no inhibitoria provocó un aumento y / o disminución en el metabolismo de TST mediado por CYP entre HLM de un solo donante en función del tamaño y la modificación del cambio de superficie. ANOVA indicó que cambios significativos por tamaño de AuNP ( p <0,0001), revestimientos superficiales ( p <0,0001) y formación de PC ( p <0.0001) were observed for the production of six selected metabolites of TST in single donor HLM (HDA1, HDB2, and HDC3).

Effects of 40 nm naked and PC AuNP on the production of 2β-OH TST (A1–A3), 6β-OH TST (B1–B3), 15β-OH TST (C1–C3), 16α-OH TST (D1–D3), 16β-OH TST (E1–E3), and AD (F1–F3); in three different single donor HLM (HDA1, HDB2, and HDC3). Means followed by the same letter were not significantly different for each nanoparticle (Tukey’s honest significant difference =5%). Naked no PC, PC human plasma protein corona, HLM human liver microsomes, BPEI branched polyethylenimine, LA lipoic acid, PEG polyethylene glycol, 2β -OH TST 2β-hydroxytestosterone, 6β -OH TST 6β-hydroxytestosterone, 15β -OH TST 15β-hydroxytestosterone, 16α -OH TST 16α-hydroxytestosterone, 16β -OH TST 16β-hydroxytestosterone, AD androstenedione

Effects of 80 nm naked and PC AuNP on the production of 2β-OH TST (A1–A3), 6β-OH TST (B1–B3), 15β-OH TST (C1–C3), 16α-OH TST (D1–D3), 16β-OH TST (E1–E3), and AD (F1–F3); in three different single donor HLM (HDA1, HDB2, and HDC3). Means followed by the same letter were not significantly different for each nanoparticle (Tukey’s honest significant difference =5%). Naked no PC, PC human plasma protein corona, HLM human liver microsomes, BPEI branched polyethylenimine, LA lipoic acid, PEG polyethylene glycol, 2β -OH TST 2β-hydroxytestosterone, 6β -OH TST 6β-hydroxytestosterone, 15β -OH TST 15β-hydroxytestosterone, 16α -OH TST 16α-hydroxytestosterone, 16β -OH TST 16β-hydroxytestosterone, AD androstenedione
All 40 nm naked and PC AuNP decreased both 2β-OH TST and 6β-OH TST production in HDA1 and HDC3 except for PC PEG-AuNP in the former and naked LC-AuNP in the latter, whereas in HDB2, only PC AuNP potentiated inhibition of their productions, irrespective of surface coatings (Fig. 8(A1–B3)). The 40 nm naked LA-AuNP was an inhibitor for 15β-OH TST production in HDA1; for HDB2 PC PEG-AuNP; and for HDC3 naked BPEI- and PEG-AuNP and PC LA- and PC PEG-AuNP (Fig. 8(C1–C3)). All 40 nm naked AuNP were an activator for 16α-OH TST production in HDA1 but PC attenuated it except for PC BPEI-AuNP which potentiated its inhibition (Fig. 8(D1)). All 40 nm naked and PC AuNP did not influence the production of 16β-OH production within individuals (Fig. 8(E1–E3)). AD production was modulated by the 40 nm naked and PC AuNP with varying degrees of inhibition except for PC PEG-AuNP which served an activator in HDC3 (Fig. 8(F1–F3)).
The 80 nm naked and PC AuNP-mediated inhibition for 2β-OH TST production was observed within individuals except for 80 nm naked and PC PEG-AuNP which served as the activators in HDB2 and HDC3, respectively (Fig. 9(A1–A3)). These results were not consistent with all naked and PC 40 nm AuNP-mediated inhibition for 2β-OH TST, irrespective of surface coatings (Fig. 8(A1–A3)). For 6β-OH TST, the 80 nm naked AuNP were the activators except for LA-AuNP but PC potentiated its inhibition in HDB2 (Fig. 9(B2)), which was similar to the 40 nm naked and PC AuNP-mediated inhibition and/or activation, respectively (Fig. 8(B2)). For 15β-OH TST, 80 nm naked BPEI- and PEG-AuNP and PC PEG-AuNP were the activators in HDB2 and in HDC3, respectively (Fig. 9(C2 and C3)). The 80 nm naked AuNP increased 16α-OH TST production in HDA1, irrespective of surface coatings but PC attenuated it except for PC LA-AuNP which was an inhibitor (Fig. 9(D1)). This is similar to the 40 nm naked and PC AuNP-mediated activation and attenuation for 16α-OH TST production in HDA1 (Fig. 8(D1)). All 80 nm naked and PC were not inhibitory to 16β-OH TST production within all individuals, irrespective of surface coatings (Fig. 9(E1–E3)). These results were consistent with the 40 nm naked and PC-mediated its production within individuals (Fig. 8(E1–E3)). For AD production, the 80 nm naked BPEI- and PEG-AuNP were the inhibitors but PC attenuated and vice versa with naked and PC LA-AuNP in HDA1 (Fig. 9(F1)). The 80 nm naked and PC AuNP decreased its production in HDC3 except for PC PEG-AuNP, which was an activator (Fig. 9(F3)). This study strongly suggests that AuNP interaction with CYP enzymes in HLM cause a decrease and/or increase in TST conversion to hydroxylated and dealkylated metabolites within individuals and the presence of PC played the inhibitive or protective role. In vivo study reported that the male CD-1 mice orally administrated with ketoconazole, a noncompetitive CYP3A4 inhibitor showed that a decrease in serum TST level, gonadal TST secretion, and hepatic TST hydroxylation activity that included 6β-OH TST, 15α-OH TST, 15β-OH TST, and 16β-OH TST [34]. In vitro studies with human hepatocyte, C3A cell line, HepG2 cell line, HLM, and recombinant CYP enzymes suggested that AuNP modulated the activity of various CYP enzymes that included CYP1A2, 2C9, 2C19, 2D6, 2E1, and 3A4 [6, 7, 20, 21]. PC and human serum albumin corona mitigated an inhibitory effect of BPEI- and LA-AuNP on CYP1A2, 2C9, and 3A4 enzyme activity in human hepatocytes and C3A cell line [6, 7]. That being said, it may be rational to propose that AuNP interference with CYP enzymes relates individual susceptibility to unexpected toxicological effects that may result in an altered circulating TST level tied to endocrine disrupting substance and/or drug-drug interaction sharing the same CYP enzymes [35].
Conclusiones
These studies exhibit that AuNP interaction with PC definitely modulate CYP-dependent metabolism of TST in HLM derived from a large donor pool that better represents the average American population. The 40 nm naked (no PC) AuNP and to a lesser degree 80 nm naked AuNP inhibited TST hydroxylation but activated TST dealkylation at high concentration. Cationic BPEI-AuNP withheld the production of 6β-OH TST and 15β-OH TST in pooled HLM but the presence of a more biologically relevant PC alleviated their adverse effects as function of size and surface charge modification. In most cases, the 40 and 80 nm naked and PC AuNP are essentially inhibitory to TST metabolism in single donor HLM in a surface chemistry-dependent manner at the noninhibitory concentration. In addition, PC PEG-AuNP caused an activation of AD production in HDC3, irrespective of size. These results may indicate that individual variations in AuNP-mediated TST metabolism could be a factor for their toxicity and could be utilized to identify vulnerable subgroup to TST-disrupting NP.
Disponibilidad de datos y materiales
All data generated or analyzed during this study are included in this article and its supplementary information file.
Abreviaturas
- 11β-OH TST:
-
11β-hydroxytestosterone
- 15β-OH TST1:
-
5β-hydroxytestosterone
- 16α-OH TST:
-
16α-hydroxytestosterone
- 16β-OH TST:
-
16β-hydroxytestosterone.
- 2α-OH TST:
-
2α-hydroxytestosterone
- 2β-OH TST:
-
2β-hydroxytestosterone
- 6α-OH TST:
-
6α-hydroxytestosterone
- 6β-OH TST:
-
6β-hydroxytestosterone
- AD:
-
Androstenedione
- AgNP:
-
Nanopartículas de plata
- ANOVA:
-
One-way analysis of variance
- AuNP:
-
Nanopartículas de oro
- BPEI:
-
Branched polyethylenimine
- CFS:
-
Chlorpyrifos
- CYP:
-
Cytochrome P450
- DEET:
-
Diethyltoluamide
- DH :
-
Hydrodynamic diameters
- DI:
-
Agua desionizada
- DLS:
-
Dispersión de luz dinámica
- EC50 :
-
Half maximal activation concentration
- EDC/NHS:
-
1-Ethyl-3-(3-dimethylaminopropyl) carbodiimide/N-hydroxysuccinimide
- ESI + :
-
Electrospray positive
- HLM:
-
Human liver microsomes
- HSD:
-
Tukey’s honest significant difference
- IC50 :
-
Half maximal inhibitory concentration
- ISTD:
-
Internal standard
- LA:
-
Lipoic acid
- LC-MS/MS:
-
Liquid chromatography-mass spectrometry
- LOD:
-
Límite de detección
- LOQ:
-
Limit of quantitation
- MRM:
-
Multiple reaction monitoring
- NADP:
-
Nicotinamide adenine dinucleotide phosphate
- NADPH:
-
Reduced NADP
- naked:
-
No PC
- NP:
-
Nanopartículas
- PBS:
-
Solución salina tamponada con fosfato
- PC:
-
Human plasma protein corona
- PDI:
-
Índice de polidispersidad
- PEG:
-
Polietilenglicol
- pHLM:
-
Pooled human liver microsomes
- QC:
-
Quality control
- SWCNT:
-
Single-walled carbon nanotube
- TEM:
-
Microscopía electrónica de transmisión
- TFF:
-
Tangential flow filtration
- TiO 2 :
-
Titanium dioxide
- TST:
-
Testosterone
- UPLC TQD:
-
Ultra performance liquid chromatography system with Triple quadrupole Detector
Nanomateriales
- Nanopartículas semiconductoras
- Nanopartículas plasmónicas
- Nanopartículas de oro para sensores de quimioterapia
- Nanopartículas de oro multifuncionales para aplicaciones terapéuticas y diagnósticas mejoradas:una revisión
- Nanobiosensor de oro basado en la resonancia de plasmón superficial localizado es capaz de diagnosticar la brucelosis humana, presentando un método rápido y asequible
- La posible toxicidad hepática, cerebral y embrionaria de las nanopartículas de dióxido de titanio en ratones
- Estabilidad mejorada de nanopartículas magnéticas de oro con poli (ácido 4-estirenosulfónico-ácido co-maleico):propiedades ópticas personalizadas para la detección de proteínas
- Poliglicerol hiperramificado modificado como dispersante para el control del tamaño y la estabilización de nanopartículas de oro en hidrocarburos
- Promoción del crecimiento celular SH-SY5Y mediante nanopartículas de oro modificadas con 6-mercaptopurina y un péptido penetrante de neuronas
- Saponinas Platycodon de Platycodi Radix (Platycodon grandiflorum) para la síntesis verde de nanopartículas de oro y plata
- Tatuaje hecho con nanopartículas de oro revoluciona el diagnóstico médico