


Un nuevo mecanismo subyacente a la formación de raíces laterales de tomate activada por nanotubos de carbono de paredes múltiples:la participación del óxido nítrico
Resumen
Numerosos estudios revelaron que los nanotubos de carbono de paredes múltiples (MWCNT) son tóxicos para las plantas. Sin embargo, si los MWCNT influyen en la formación de raíces laterales (LR), que es un componente importante de la adaptabilidad del sistema de raíces a diversas señales ambientales, sigue siendo controvertido. En este informe, encontramos que los MWCNT podrían entrar en las raíces de las plántulas de tomate. La administración con MWCNT promovió la formación de LR en tomate de una forma aproximadamente dependiente de la dosis. La producción endógena de óxido nítrico (NO) fue desencadenada por MWCNT, confirmada por el método de reactivo de Greiss, resonancia paramagnética electrónica (EPR) y microscopía confocal de barrido láser (LSCM), junto con el eliminador de NO. Existe una relación de causa-efecto entre MWCNT y NO en la inducción del desarrollo de LR, ya que la síntesis de NO activada por MWCNT y la formación de LR fueron obviamente bloqueadas por la eliminación de NO endógeno con su eliminador. La actividad de la enzima nitrato reductasa (NR) generadora de NO aumentó en respuesta a los MWCNT. La inhibición de Tungstato de NR no solo afectó la producción de NO, sino que también abolió la formación de LR desencadenada por MWCNT. La adición de N G -nitro-l-arginina metil éster (l-NAME), un inhibidor de la enzima similar a la óxido nítrico sintasa (NOS) de mamíferos, no influyó en la formación de LR. Colectivamente, propusimos que el NO podría actuar como una molécula de señalización aguas abajo en el control de MWCNT del desarrollo de LR, al menos parcialmente a través de NR.
Introducción
Existen muchas aplicaciones biológicas y biomédicas de los nanotubos de carbono [1, 2]. Debido a la capacidad única de penetrar fácilmente las membranas celulares, la bioseguridad de los nanotubos de carbono es siempre un tema de debate [3, 4]. Mientras tanto, dado que la producción y el uso de nanotubos de carbono crecen rápidamente, es importante caracterizar los mecanismos detallados de su citotoxicidad en seres humanos y mamíferos, y recientemente en plantas [3,4,5,6,7,8,9]. Es bien sabido que las plantas y sus comunidades son muy importantes para los seres humanos y el medio ambiente, y también corren el riesgo de exposición a los nanotubos de carbono, debido a la acumulación en el suelo a través de la aplicación de fertilizantes biosólidos [6, 10, 11]. Como miembros importantes de los nanotubos de carbono, se ha investigado ampliamente la toxicidad de los nanotubos de carbono de paredes múltiples (MWCNT), que consisten en múltiples capas enrolladas de grafeno. Los estudios en mamíferos revelaron que la exposición con MWCNT y nanotubos de carbono de pared simple inducía daño oxidativo y activación de NF-κB en queratinocitos humanos y células A549 [9, 12]. Los MWCNT y los nanotubos de carbono de pared simple pueden fusionarse con la membrana plasmática, causando así daño celular a través de la peroxidación lipídica y el estrés oxidativo [9, 11, 13, 14]. Se observaron citotoxicidad y estrés oxidativo desencadenado por MWCNT, así como respuestas inflamatorias modestas, en células endoteliales de la vena umbilical humana [15]. Un estudio anterior sugirió que la toxicidad primaria de los MWCNT en la espinaca roja se derivaba principalmente de la sobreproducción de especies reactivas de oxígeno (ROS), y los efectos tóxicos podrían revertirse con el ácido ascórbico suplementado [7]. En este sentido, los MWCNT se consideran un nuevo factor de estrés para los organismos, ya sea en animales o en plantas.
La formación de raíces laterales (LR), un determinante importante de la arquitectura de las raíces, se ha considerado un indicador de la respuesta adaptativa a diversas tensiones [16]. En plantas superiores, la formación de LR está influenciada por fitohormonas y una amplia gama de señales ambientales, incluida la disponibilidad de agua, nutrientes y estrés abiótico, como hipoxia y estrés por metales pesados [17,18,19]. Mientras tanto, una amplia evidencia confirmó que la formación de LR no solo actúa como un soporte físico, sino que también contribuye a la absorción de agua y nutrientes para el crecimiento y desarrollo de las plantas [19,20,21]. Diferentes pistas ambientales podrían desencadenar varios fenotipos específicos de respuesta morfogénica inducida por estrés (SIMR), incluida la promoción de la formación de LR y la inhibición de la elongación de la raíz [17]. La regulación de la formación de LR también está estrictamente controlada por fitohormonas, como la auxina, y se sugirió la activación de genes reguladores del ciclo celular en respuesta a la auxina [19, 22]. Mientras tanto, se confirmó la participación de algunas moléculas pequeñas en la organogénesis de la raíz desencadenada por auxinas en plantas de pepino, tomate, soja y colza [23,24,25,26,27]. Estas pequeñas moléculas incluyen peróxido de hidrógeno (H 2 O 2 ), óxido nítrico (NO), monóxido de carbono (CO) e hidrógeno gaseoso (H 2 ).
Entre estos, se ha demostrado que el NO, un gas de radicales libres, tiene múltiples funciones fisiológicas en las plantas [28, 29]. Además de la mejora de la adaptación de la planta contra el estrés, las funciones del NO incluyen la promoción del desarrollo del vello radicular, el enraizamiento adventicio y la formación de raíces laterales [30,31,32,33], aunque los recursos enzimáticos de la biosíntesis de NO en aquellos los procesos mencionados siguen siendo esquivos. En los animales, la síntesis de NO a partir de l-arginina es catalizada por la enzima óxido nítrico sintasa (NOS) que contiene hemo [34]. Aunque no se han identificado genes que codifican enzimas NOS en plantas, la actividad similar a la NOS en mamíferos se detecta ampliamente [35, 36], y los inhibidores de la NOS en mamíferos, como N G -clorhidrato de éster metílico de nitro-l-arginina (l-NAME), puede inhibir la generación de NO en plantas [25, 33, 36,37,38,39]. Es importante destacar que una amplia evidencia genética reveló que el NO puede ser producido por la nitrato reductasa (NR), una enzima conocida responsable del metabolismo del nitrógeno en las plantas [28]. La implicación de la producción de NO mediada por NR en el cierre de los estomas y la aclimatación al frío se ha demostrado genéticamente [37, 38]. Nuestro estudio anterior mostró que la síntesis de NO dependiente de NR está involucrada en la formación de raíces laterales mediada por gas hidrógeno inducida por auxinas [39].
Hasta ahora, se informaron diferentes respuestas en la formación, promoción o inhibición de LR, respectivamente, en varias especies de plantas cuando se complementaron con nanomateriales, incluidos MWCNT [40,41,42,43], nanopartículas de oro (Au NP, [44]), óxido de zinc nanopartículas (ZnO NP [45, 46];), nanopartículas de dióxido de titanio (TiO 2 NP [46];) y óxido de grafeno (GO [47,48,49];) (Tabla 1), y ningún estudio ha proporcionado una prueba definitiva del papel del NO en las respuestas anteriores. En este estudio, la detección de NO endógeno por el método de reactivo de Greiss, microscopía confocal de barrido láser (LSCM) y análisis de resonancia paramagnética electrónica (EPR) revelaron que el nivel de NO aumentó en plántulas de tomate tratadas con MWCNT. Posteriormente, se observó la formación de LR. Además, estudiamos la participación de NO en la formación de LR desencadenada por MWCNT, mediante la manipulación de los niveles de NO endógenos utilizando un eliminador de NO y antagonistas que inhiben la NR y la actividad de NOS similar a la de los mamíferos. Experimentos posteriores revelaron que el NO dependiente de NR podría ser, al menos parcialmente, esencial para la formación de LR en respuesta a los MWCNT. Por lo tanto, este trabajo abre una nueva ventana para comprender los efectos biológicos de los nanomateriales en las plantas.
Materiales y métodos
Productos químicos
A menos que se indique lo contrario, todos los demás productos químicos se obtuvieron de Sigma-Aldrich (St Louis, MO, EE. UU.). Los MWCNT, adquiridos de Sigma-Aldrich, se caracterizaron como se describió anteriormente [50]. El diámetro exterior, el diámetro interno y la longitud de los MWCNT fueron de 6 a 12 nm, de 2,5 a 5 nm y de 1 a 9 μm, respectivamente. Después del tratamiento de sonicación, la suspensión coloidal homogeneizada obtenida se esterilizó y utilizó.
Se obtuvieron otras nanopartículas de carbono de Nanjing XFNANO Materials Tech Co., Ltd., incluidos los nanotubos de carbono de pared simple (SWCNT, XFS22; pureza> 95%, diámetro 1 a 2 nm, longitud 5 a 30 μm, área de superficie especial> 1075 m 2 / g), grafeno (XF001W; pureza ~ 99%, diámetro 0,5–5 μm, grosor ~ 0,8 nm, relación de una sola capa ~ 80%, área de superficie BET 500 ~ 1000 m 2 /gramo; resistividad eléctrica ≦ 0,30 Ω.cm) y carbón activo (AC, XFP06; pureza> 95%, tamaño de partícula 5 ± 1 μm, volumen de poro 1–1,2 cm 3 / g, apertura de 2,0 a 2,2 nm, área de superficie especial ~ 1500-1700 m 2 / g).
Además, se utilizó nitroprusiato de sodio (SNP) como compuesto liberador de NO [30,31,32,33]. La sal de potasio de 2- (4-carboxifenil) -4,4,5,5-tetrametilimidazolin-1-oxil-3-óxido (cPTIO) se consideró como un eliminador de NO [51,52,53,54]. Tungstato (Tg; un inhibidor de NR [28, 33, 37, 55, 56, 57];) y N G También se aplicó hidrocloruro de éster metílico de nitro-L-arginina (NAME; un inhibidor de la enzima similar a la NOS de mamíferos [25, 33, 36,37,38,39];). En este estudio, las concentraciones de los productos químicos anteriores se determinaron en los experimentos piloto, a partir de los cuales se observaron las respuestas significativas.
Material vegetal y condiciones de crecimiento y determinación de la formación LR
Tomate ( Solanum lycopersicum L.) semillas “Jiangshu 14” fueron amablemente suministradas por los Institutos Agrícolas de Jiangsu, Nanjing, provincia de Jiangsu, China. Se hicieron germinar semillas seleccionadas de idéntico tamaño en agua destilada a 25 ± 1 ° C en la oscuridad durante 3 días. Las plántulas idénticas seleccionadas con radículas de 2 a 3 mm se transfirieron luego a soluciones de tratamiento de 6 ml que contenían las concentraciones indicadas de MWCNT, ácido 1-naftilacético (NAA; una auxina bien conocida) 200 nM, SNP 0,1 mM, cPTIO 0,2 mM, 20 Tungstato μM (Tg), NOMBRE 0.2 mM y otras nanopartículas de carbono, incluidos nanotubos de carbono de pared simple (SWCNT) de 5 mg / ml, grafeno y carbón activo (AC), solos o en combinación durante los puntos de tiempo indicados. Las plántulas se cultivaron en una incubadora de iluminación (25 ± 1 ° C) con una intensidad de luz de 200 μmol m −2 s −1 a las 14/10 h fotoperiodo (claro / oscuro).
Después de los tratamientos, se tomaron fotografías y se determinó el número y la longitud de la raíz lateral emergida (> 1 mm) por plántula utilizando el software Image J (http://rsb.info.nih.gov/ij/) [39 , 58]. Como se describió anteriormente, solo los segmentos inducibles por raíces laterales se utilizaron para el análisis posterior.
Imágenes de la distribución de MWCNT por microscopía electrónica de transmisión
La distribución de MWCNT en la raíz de plántula de tomate se caracterizó mediante microscopía electrónica de transmisión (TEM; JEOL, JEM-200CX, Tokio, Japón). La preparación de la muestra para el análisis TEM se realizó de acuerdo con el protocolo anterior [59].
Obtención de imágenes de NO endógeno mediante microscopio confocal de barrido láser
La formación de imágenes de NO se llevó a cabo usando una sonda fluorescente de NO bastante específica 4-amino-5-metilamino-2 ', 7'-diacetato de difluorofluoresceína (DAF-FM DA). Después de que la sonda se lavó a fondo, las imágenes se obtuvieron utilizando el microscopio confocal Zeiss LSM 710 (Carl Zeiss, Oberkochen, Alemania, excitación a 488 nm, emisión a 500–530 nm para análisis de NO). En nuestro experimento, se seleccionaron al azar 20 muestras individuales y se midieron por tratamiento. Las fotografías son representativas de resultados idénticos.
SIN contenido determinado por el ensayo de reactivo de Griess
De acuerdo con los métodos descritos anteriormente [50], el contenido de NO se determinó con el ensayo de reactivo de Griess. Es importante destacar que para escapar de la interferencia causada por los contenidos concentrados de nitrato y nitrito en las plantas, las muestras idénticas preincubadas en cPTIO 200 μM (el eliminador de NO) durante 30 min se consideraron muestras en blanco. Después de la adición de reactivo de Griess durante 30 minutos, se registró la absorbancia a 540 nm y se determinó el contenido de NO mediante comparación con una curva estándar de NaNO 2 .
Determinación de NO con resonancia paramagnética electrónica (EPR)
De acuerdo con nuestros métodos anteriores [39, 55, 60], se llevó a cabo la determinación del nivel de NO mediante resonancia paramagnética electrónica (EPR). La capa de disolvente orgánico se utilizó para determinar el NO en un espectrómetro Bruker A300 (Bruker Instrument, Karlsruhe, Alemania) en las siguientes condiciones:temperatura ambiente; frecuencia de microondas, 9,85 GHz; potencia de microondas, 63,49 mW; y frecuencia de modulación, 100,00 kHz.
Determinación de la actividad de nitrato reductasa (NR)
La actividad NR se detectó espectrofotométricamente a 540 nm de acuerdo con el método anterior [57]. El nitrito producido se determinó espectrofotométricamente a 540 nm mediante la adición de 1 ml de sulfenilamida al 1% (p / v) en HCl 3 M junto con 1 ml de n- (1-naftil) -etilendiamina al 0,02% (v / v).
Análisis estadístico
Cuando se indique, los resultados se expresaron como los valores medios ± EE de tres experimentos independientes con tres réplicas biológicas para cada uno. El análisis estadístico se realizó con el software SPSS Statistics 17.0. Para el análisis estadístico, la prueba múltiple de Duncan ( p <0.05) se eligió según corresponda.
Resultados
Los MWCNT no solo entran en las células raíz, sino que también promueven la formación de LR
La formación de LR es un determinante importante de la arquitectura de los sistemas de raíces. Para investigar el efecto de los MWCNT en la formación de LR, se incubaron plántulas de tomate de 3 días con un rango de concentraciones de MWCNT (0,05, 0,5, 5 y 50 mg / ml) durante 3 días. La aplicación de ácido 1-naftilacético (NAA) se consideró un control positivo. En nuestro experimento, tanto el número de LR como la longitud se determinaron como dos parámetros de la formación de LR. Como se muestra en la Fig. 1, en comparación con las muestras de control, los MWCNT exógenos indujeron significativamente la formación de LR de una manera aproximadamente dependiente de la dosis, con un efecto máximo en 5 y 50 mg / ml. Se observó una respuesta inducible similar cuando se administró NAA 200 nM. Teniendo en cuenta el costo de los MWCNT y la respuesta inducible en la formación de LR, se aplicó MWCNT de 5 mg / ml en los siguientes experimentos.
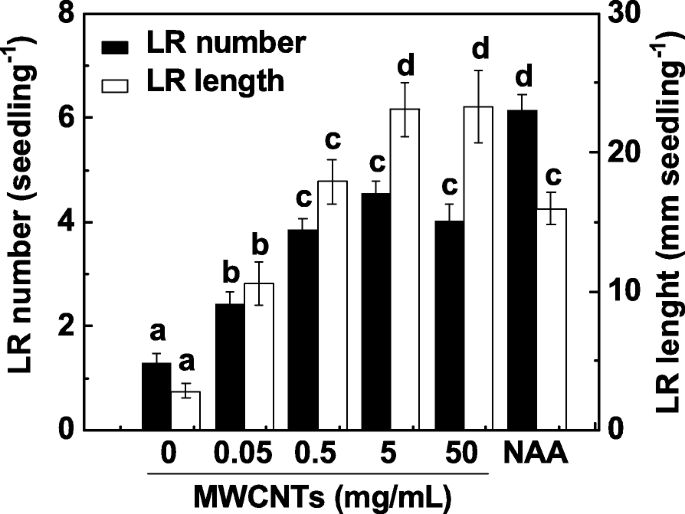
La formación de LR de tomate inducida por MWCNT fue de una manera aproximadamente dependiente de la dosis. Las plántulas de tomate de tres días se trataron con NAA 200 nM y las concentraciones indicadas de MWCNT, respectivamente. Luego se determinó el número y la longitud de la raíz lateral emergida (> 1 mm) por plántula después de 3 días de tratamiento. Había 30 (10 × 3) plantas en tres réplicas biológicas, y los experimentos se realizaron 3 veces. Los datos son las medias ± SE. Dentro de cada conjunto de experimentos, las barras indicadas por la misma letra no difirieron significativamente en p <0.05 nivel según la prueba múltiple de Duncan
Para validar la función específica de MWCNT en la inducción de la formación de LR, investigamos más a fondo si las otras alotropías de MWCNT también tienen tales efectos inducibles. Como se muestra en la Fig. 2a, todos estos nanomateriales de carbono exhibieron efectos tóxicos sobre el crecimiento de los brotes (datos no mostrados). Curiosamente, la aplicación de MWCNT, nanotubos de carbono de pared simple (SWCNT), grafeno y carbón activo con concentración idéntica (5 mg / ml) podría resultar de manera diferencial en aumentos en el número y la longitud de LR, en comparación con las plantas de control libres de químicos. (Figura 2b). Entre estos productos químicos, la respuesta inducible máxima se descubrió en plántulas de tomate incubadas con MWCNT.
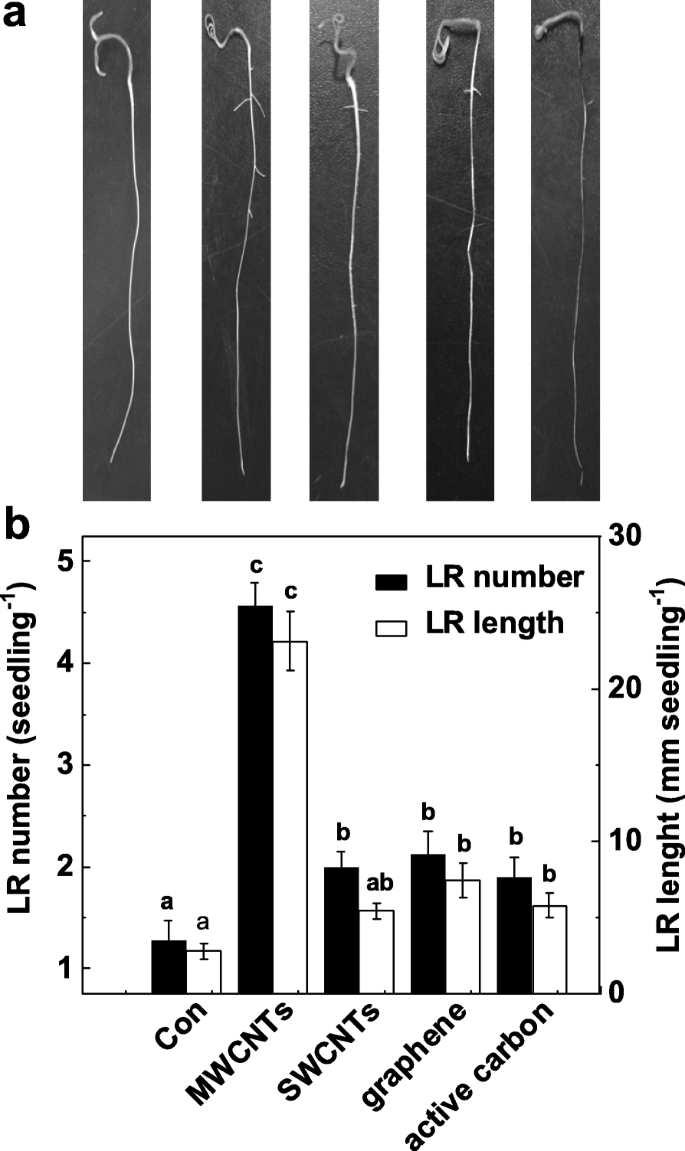
Cambios en la formación de raíces laterales en respuesta a diferentes nanopartículas de carbono. Las plántulas de tomate de tres días se trataron con agua destilada (Con), 5 mg / mL de MWCNT, nanotubos de carbono de pared simple (SWCNT), grafeno y carbón activo (AC), respectivamente, durante otros 3 días. un Luego se tomaron fotografías representativas. b También se determinó el número y la longitud de la raíz lateral emergida (> 1 mm) por plántula. Barra de escala =50 mm. Había 30 (10 × 3) plantas en tres réplicas biológicas, y los experimentos se realizaron 3 veces. Los datos son las medias ± SE. Dentro de cada conjunto de experimentos, las barras indicadas por la misma letra no difirieron significativamente en p <0.05 nivel según la prueba múltiple de Duncan
Con la ayuda de la microscopía electrónica de transmisión (TEM), la distribución de MWCNT se puede evaluar fácilmente. Los resultados mostrados en la Fig. 3 revelaron que los MWCNT, cuando se aplicaban exógenamente, podían ser absorbidos por las plántulas de tomate, y se encontró que la distribución de los MWCNT estaba en las células de la raíz. Este resultado se puede entender, ya que las raíces de las plántulas se cultivan directamente en una solución líquida que contiene MWCNT.
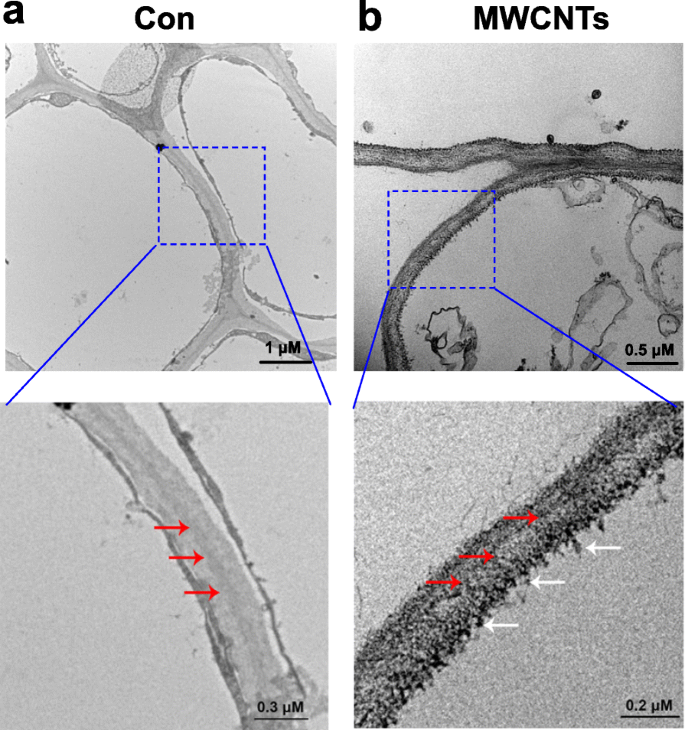
Distribución de MWCNT en raíces de tomate. Imágenes TEM de plántulas de tomate de 3 días tratadas con agua destilada (Con; a ) o MWCNT de 5 mg / ml ( b ) durante 1 día. La flecha roja indica la pared celular, mientras que la flecha blanca indica MWCNT
La síntesis de NO inducida por MWCNT y, posteriormente, la formación de LR fueron sensibles a cPTIO, un eliminador de NO
Para investigar si el NO también está implicado en la formación de LR inducida por MWCNT, se evaluó la función de NO en la formación de LR provocada por los MWCNT manipulando los niveles de NO endógenos usando el compuesto liberador de NO y el eliminador. De forma similar a los resultados anteriores [31], la administración de nitroprusiato de sodio (SNP) podría provocar la inducción de la formación de LR, y se observó una respuesta aditiva cuando se aplicaron conjuntamente SNP y MWCNT (Fig. 4). Cuando se añadió sal potásica de 2- (4-carboxifenil) -4,4,5,5-tetrametilimidazolin-1-oxil-3-óxido (cPTIO; un eliminador de NO), las respuestas de promoción en la formación de LR causadas por MWCNT fueron significativamente dañado. Por sí solo, cPTIO podría inhibir el desarrollo de LR, en comparación con el control sin químicos, lo que indica el importante papel del NO endógeno en la organogénesis de la raíz.
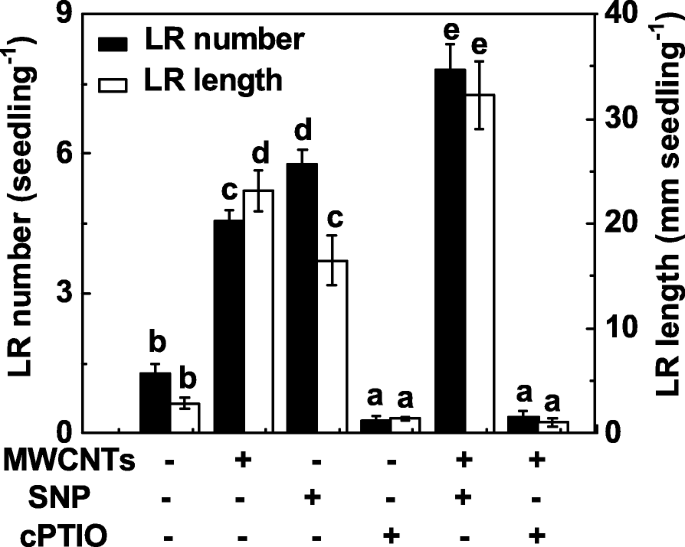
La formación de LR inducida por MWCNT fue sensible a la eliminación de NO endógeno con cPTIO, su eliminador. Las plántulas de tomate de tres días se trataron con agua destilada, MWCNT 5 mg / ml, SNP 0,1 mM, cPTIO 0,2 mM, solos o en combinación durante 3 días. Posteriormente, se determinó el número y la longitud de la raíz lateral emergida (> 1 mm) por plántula. Había 30 (10 × 3) plantas en tres réplicas biológicas, y los experimentos se realizaron 3 veces. Los datos son las medias ± SE. Dentro de cada conjunto de experimentos, las barras indicadas por la misma letra no difirieron significativamente en p <0.05 nivel según la prueba múltiple de Duncan
Con el fin de evaluar adicionalmente el importante papel del NO endógeno en la respuesta de MWCNT, en primer lugar se detectó un curso temporal de la producción de NO in vivo con el método del reactivo de Greiss. Durante la determinación anterior, el filtrado idéntico pretratado con cPTIO se consideró como un blanco para obtener resultados precisos. Se observó que la producción de NO en las raíces de las plántulas de tomate aumentó drásticamente hasta 24 h después del tratamiento con MWCNT y luego se recupera a los niveles iniciales (48 h; Fig. 5a). Por encima del nivel máximo de NO endógeno desencadenado por MWCNT durante 24 h fue obviamente abolido por cPTIO, un eliminador de NO, lo que sugiere el papel específico de NO.
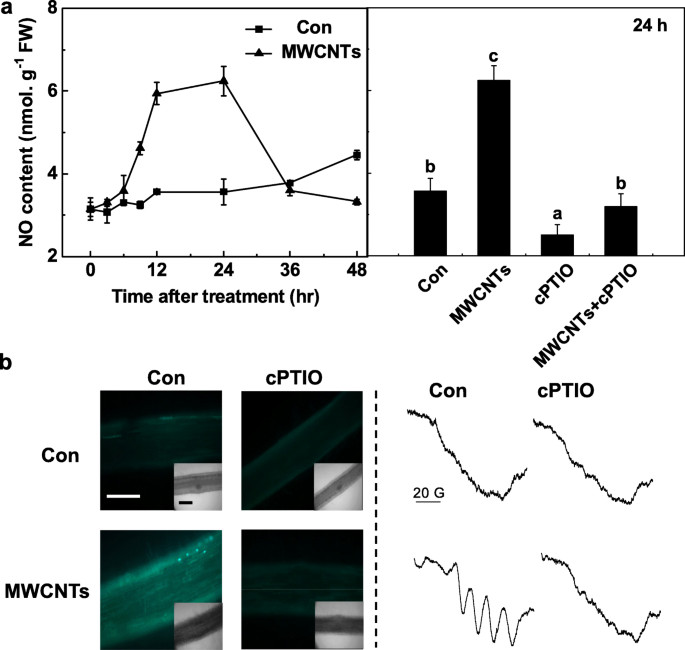
La producción de NO inducida por MWCNT fue bloqueada por cPTIO, el eliminador de NO. Se trataron plántulas de tomate de tres días con agua destilada y MWCNT de 5 mg / ml con o sin cPTIO 0,2 mM, respectivamente. un Los cambios en la actividad de NR (izquierda) y la producción de NO (derecha) se determinaron mediante el método de reactivo de Greiss. b Después de un tratamiento de 24 h, la señal de NO se analizó mediante LSCM (izquierda) y EPR (derecha). Barra de escala =0,1 mm. Los datos son las medias ± SE. Las barras indicadas por la misma letra no difieren significativamente en p <0.05 nivel según la prueba múltiple de Ducan
Para confirmar los resultados anteriores, se adoptaron tanto LSCM como ESR. En primer lugar, se controlaron los cambios en los niveles de NO endógeno en las raíces de las plántulas de tomate marcando NO usando la sonda fluorescente DAF-FM DA permeable a las células y bastante específica de NO y obteniendo imágenes con LSCM. De manera similar a los resultados anteriores (Fig. 5a), en presencia de cPTIO, la intensidad de fluorescencia dependiente de DAF-FM aumentada desencadenada por MWCNT se abolió en gran medida (Fig. 5b). Estos resultados implicaron que la fluorescencia desencadenada por DAF-FM está relacionada con los niveles de NO endógeno en las raíces de las plántulas de tomate.
La producción de NO inducida por MWCNT se confirmó mediante espectroscopía EPR. Como se esperaba, las raíces de las plántulas tratadas durante 24 h con MWCNT presentaron el típico triplete de estructura hiperfina del complejo NO. Sin embargo, la adición de cPTIO abolió la señal anterior, lo que indica que la exposición a MWCNT dio como resultado una fuerte producción de NO (Fig. 5b). En conjunto, estos datos sugirieron que la síntesis de NO podría ser necesaria para la formación de LR desencadenada por MWCNT en plántulas de tomate.
NR podría ser responsable de la producción de NO inducida por MWCNT y, posteriormente, de la formación de LR
Dado que la NR y la NOS similar a la de los mamíferos son dos enzimas principales relacionadas con la síntesis de NO en las plantas, en el experimento posterior se aplicaron tanto tungstato (un inhibidor de NR) como NAME (un inhibidor de la NOS de mamíferos). Aquí, el tratamiento con tungstato suprimió sustancialmente la promoción de la formación de LR en raíces de plántulas de tomate tratadas con MWCNT (Fig. 6). Comparativamente, la inducción de la formación de LR desencadenada por los MWCNT no fue fuertemente inhibida por la adición de NAME, lo que indica que la NOS similar a los mamíferos podría no ser la enzima sintética de NO diana responsable de la producción de NO provocada por los MWCNT. También se observó que se observó una disminución leve pero no significativa en la formación de LR en plántulas de tomate cuando se aplicó por separado tungstato o NAME.
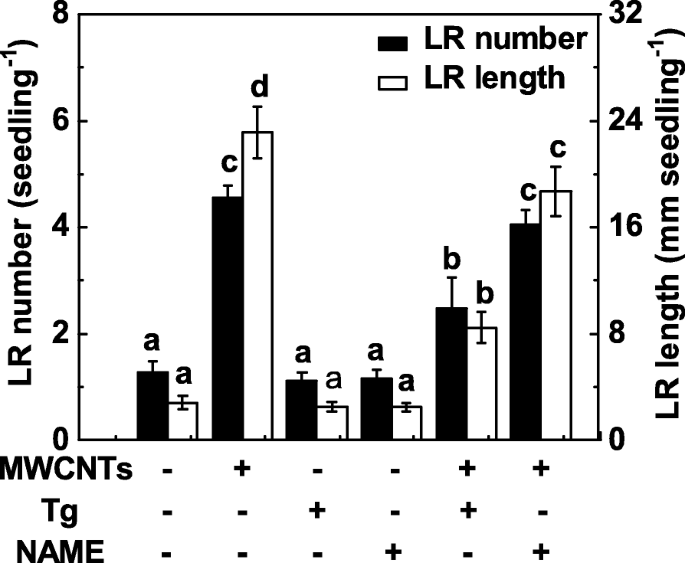
Cambios en la formación de LR en respuesta a MWCNT y dos inhibidores de la síntesis de NO. Las plántulas de tomate de tres días se trataron con MWCNT 5 mg / mL, tungstato (Tg) 20 μM, NAME 0,2 mM, solo o en combinación durante 3 días. Posteriormente, se determinó el número y la longitud de la raíz lateral emergida (> 1 mm) por plántula. Había 30 (10 × 3) plantas en tres réplicas biológicas, y los experimentos se realizaron 3 veces. Los datos son las medias ± SE. Dentro de cada conjunto de experimentos, las barras indicadas por la misma letra no difirieron significativamente en p <0.05 nivel según la prueba múltiple de Duncan
El papel de NR en la formación de LR desencadenada por MWCNT se examinó más a fondo mediante el seguimiento de la producción de NO en respuesta a MWCNT aplicados con o sin tungstato. En comparación con los cambios en la producción de NO endógeno (Fig. 5a), el análisis de la evolución temporal de la actividad NR mostró una tendencia similar (Fig. 7a), que también alcanzó un máximo a las 24 h después del tratamiento con MWCNT. Estos resultados sugirieron que el aumento inducido por MWCNT en la producción de NO puede resultar principalmente de una actividad mejorada de NR. De manera consistente, la inhibición de la producción de NO dependiente de NR por el tungstato se confirmó mediante el uso del método de reactivo de Greiss (Fig. 7b), LSCM y EPR (Archivo adicional 1:Figura S1).
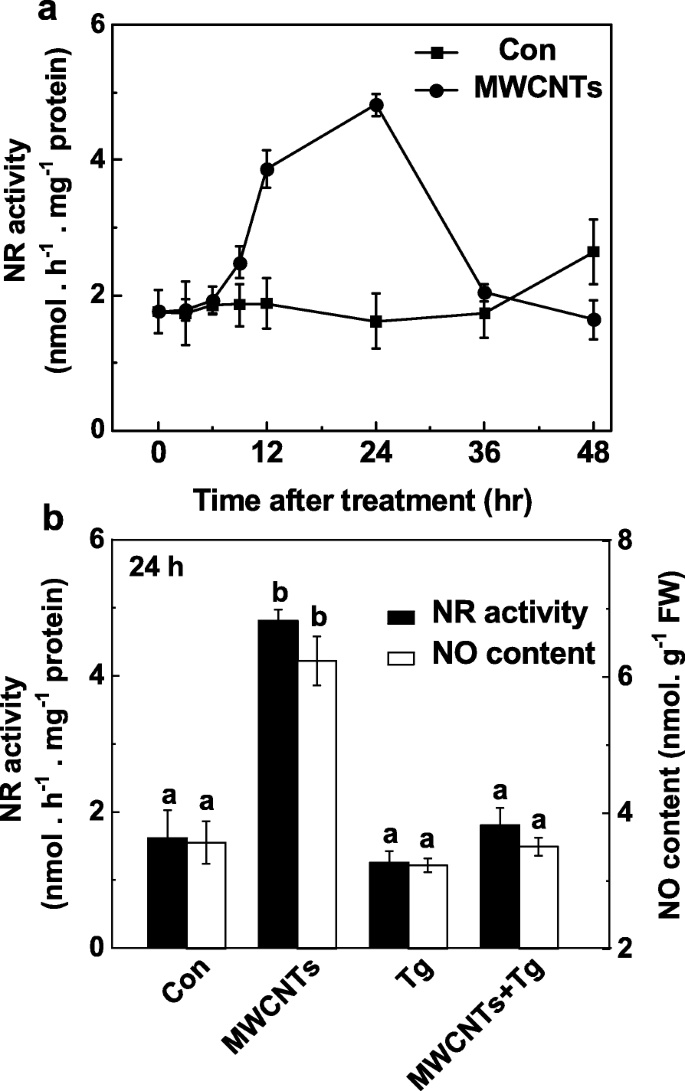
La producción de NO inducida por MWCNT fue bloqueada por tungstato, un inhibidor de NR. Las plántulas de tomate de tres días se trataron con agua destilada y MWCNT de 5 mg / ml con o sin tungstato (Tg) 20 μM. Cambios en la actividad de NR ( a ) y NO producción ( b ) determinado utilizando el método de reactivo de Greiss. Los datos son las medias ± SE. Las barras indicadas por la misma letra no difieren significativamente en p <0.05 nivel según la prueba múltiple de Ducan
Discusión
La fitotoxicidad es una consideración importante para comprender el impacto ambiental potencial de las nanopartículas [4, 7, 61,62,63]. Abundantes pruebas revelaron que los MWCNT son tóxicos para las plantas, lo que incluye inducir daño oxidativo, inhibir la germinación de las semillas, el crecimiento y el desarrollo de las raíces [11, 63, 64]. Sin embargo, al ser un fenotipo de SIMR, la ramificación de raíces a través de la formación de raíces laterales es un componente importante de la adaptabilidad del sistema de raíces a diversas señales ambientales [17]. En este trabajo, integramos análisis biológicos, farmacológicos y bioquímicos para mostrar la participación de la producción de NO mediada por NR en la formación de LR inducida por MWCNT, al menos parcialmente en nuestras condiciones experimentales. Además, la función del NO en la organogénesis de la raíz estimulada por los MWCNT enfatizó las funciones centrales de este segundo mensajero involucrado en el proceso de desarrollo de la planta y la adaptación contra el estrés [29, 30, 31, 32, 33, 37, 38].
En primer lugar, confirmamos que los MWCNT de 5 mg / ml (DO 6–12 nm) podrían entrar en los tejidos de la raíz (Fig. 3). Posteriormente, se observó la inducción de la formación de LR en tomate (Fig. 1), imitando las funciones de inducción de NAA y SNP (Fig. 4), un conocido compuesto liberador de NO [30, 31]. Se descubrieron respuestas de inducción similares en árboles resinosos [41], lechuga [42] y Arabidopsis [43] cuando se desafiaron con MWCNT (OD 6-13 nm, aproximadamente 9,5 nm y 30-40 nm, respectivamente). Por ejemplo, la aplicación con MWCNT prístino (p-MWCNT) o MWCNT funcionalizado con carboxilo (c-MWCNT) (diámetro medio de 9,5 nm) podría promover el desarrollo de LR en plántulas de lechuga [42]. Por el contrario, la inhibición de la raíz primaria e incluso la formación de LR se encontraron simultáneamente en plantas de soja cuando se sometieron a MWCNT (DO 20-70 nm [40];). Comparando con los datos en diámetro externo de MWCNTs (Tabla 1), supusimos que los efectos exhibidos por MWCNT sobre la formación de LR variaban con sus diámetros, mostrando la promoción con menor diámetro y la inhibición con mayor diámetro. Ciertamente, el mecanismo relacionado debe investigarse cuidadosamente. Se confirmó un fenómeno similar en la tolerancia a la salinidad de las plantas [50]. En combinación con los resultados anteriores, se dedujo además que la función de los nanomateriales puede variar de una especie a otra, y viceversa, los diferentes tipos de nanomateriales pueden causar diversos efectos biológicos. Sin embargo, no se pueden descartar fácilmente otros factores influyentes, como las diferentes dosis de MWCNT [48] e incluso las condiciones de crecimiento de las plantas.
En comparación con otros nanomateriales, incluidos SWCNT, grafeno y AC con una concentración idéntica, la inducción máxima en la formación de LR e incluso los efectos tóxicos sobre el crecimiento de los brotes se observaron en los MWCNT (Fig. 2). Estos podrían estar relacionados con las características físicas especiales de los MWCNT, un tipo de nanomateriales que tienen una alta conductividad eléctrica, una gran superficie específica, una alta relación de aspecto y una notable estabilidad térmica [65]. Los efectos tóxicos de los nanomateriales se han informado ampliamente en pepino, repollo, zanahoria, cebolla, etc. [66, 67].
Una amplia evidencia mostró que el NO, actuando como una molécula de señalización, puede regular una amplia gama de procesos de las plantas desde la adaptación ambiental hasta el desarrollo y el último de los cuales incluye la germinación de las semillas y la organogénesis de las raíces [29, 68,69,70,71,72,73 ]. Nuestro experimento posterior reveló que el NO puede estar implicado en la formación de LR inducida por MWCNT. Aunque se han aplicado varios métodos para obtener imágenes de la producción de NO en células vegetales, las desventajas, incluida la falta de sensibilidad y la interferencia de moléculas independientes de NO, pueden existir en cada método [74]. Por lo tanto, se aplicaron en nuestras condiciones experimentales tres métodos responsables de la obtención de imágenes y la determinación de NO, incluido el método del reactivo de Greiss, LSCM y EPR, junto con la aplicación de cPTIO, un eliminador de NO. Mediante el uso de tres métodos, observamos que un aumento de la producción de NO endógeno inducida por MWCNT en plántulas de tomate fue abolido por cPTIO (Fig. 5), un eliminador de NO [30, 31, 32]. Es importante destacar que este proceso se correlacionó con la respuesta biológica del desarrollo de LR inducido por MWCNT, que se bloqueó severamente cuando se aplicó cPTIO simultáneamente (Fig. 4).
La evaluación adicional de estas respuestas y la (s) fuente (s) potencial (s) de NO inducida por MWCNT aplicados exógenamente reveló que la producción de NO y, posteriormente, la formación de LR podrían atribuirse a la actividad de NR. En las plantas, la producción de NO se genera principalmente a partir de NR y de proteínas similares a la NOS de mamíferos [28]. Sin embargo, el gen NOS de la planta todavía no está identificado [35, 75, 76], aunque algunos experimentos que utilizan los inhibidores de la enzima NOS de mamíferos proporcionaron alguna evidencia de la vía dependiente de la l-arginina en la producción de NO [36, 76]. Se ha confirmado que los NR son las fuentes más importantes de NO en las plantas [28]. Estudios anteriores demostraron que la producción de NO dependiente de NR funciona como una señal relacionada con los nitratos que participa en la regulación de la arquitectura de la raíz [32, 33]. Además, la producción de NO dependiente de NR se asoció estrechamente con la aclimatación al frío [38], la tolerancia a la salinidad [50] y el cierre estomático inducido por ácido abscísico [77]. Nuestros resultados revelaron además que el tungstato (un inhibidor de NR) obviamente alteraba la formación de LR inducida por MWCNT, especialmente en la longitud de LR (Fig. 6). Por el contrario, sólo hubo una ligera disminución en la longitud de LR, y no se observó una diferencia significativa en el número de LR cuando se usó l-NAME (un inhibidor de la NOS de mamíferos). De manera consistente, el ensayo bioquímico mostró que la actividad de NR aumentó obviamente por los MWCNT (Fig. 7a), en paralelo con los cambios en la producción de NO (Fig. 5a). Las respuestas anteriores podrían estar totalmente bloqueadas por tungstato (Fig. 7a, archivo adicional 1:Figura S1). Por tanto, dedujimos que el aumento de la producción de NO endógeno inducida por los MWCNT se atribuía principalmente a la vía NR. Ciertamente, se deben investigar más pruebas genéticas.
Conclusión
In summary, we provide evidence to show that MWCNT-induced NO production via NR might be required for tomato lateral root formation and this was summarized in Fig. 8. Importantly, above findings provide insights into the intricate molecular mechanism of MWCNTs functions in plants.

Schematic representation of the proposed MWCNT-induced tomato lateral root formation mainly via NR-dependent NO production. The role of mammalian NOS-like enzyme was preliminarily ruled out
Disponibilidad de datos y materiales
Todos los datos están completamente disponibles sin restricciones.
Abreviaturas
- CO:
-
Carbon monoxide
- cPTIO:
-
2-(4-Carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide potassium salt
- DAF-FM DA:
-
4-Amino-5-methylamino-2′,7′-difluorofluorescein diacetate
- EPR:
-
Resonancia paramagnética de electrones
- GO:
-
Óxido de grafeno
- H 2 :
-
Hydrogen gas
- H 2 O 2 :
-
Peróxido de hidrógeno
- l-NAME:
-
N G -Nitro-l-arginine methyl ester
- LR:
-
Lateral root
- LSCM:
-
Laser scanning confocal microscopy
- MWCNTs:
-
Multi-walled carbon nanotubes
- NAA:
-
1-Naphthylacetic acid
- NO:
-
Nitric oxide
- NOS:
-
Nitric oxide synthase
- NR:
-
Nitrate reductase
- ROS:
-
Especies reactivas de oxígeno
- SIMR:
-
Stress-induced morphogenic response
- SNP:
-
Sodium nitroprusside
- SWCNTs:
-
Single-walled carbon nanotubes
- TEM:
-
Microscopía electrónica de transmisión
- Tg:
-
Tungstate
Nanomateriales
- Los nanotubos de carbono superan el límite de la electrónica flexible
- Imagen de carbono oceánico en escala atómica
- Las ventajas de una prótesis de fibra de carbono
- Oxinitruro de boro y carbono como nuevo fotocatalizador sin metales
- Presentación de la estructura atómica y electrónica de las nanofibras de carbono de copa apilada
- La detección fotoelectroquímica mejorada de ácido úrico en electrodo de carbono vítreo modificado con nanopartículas de Au
- El estudio de un nuevo sistema micelar similar a un gusano mejorado con nanopartículas
- Los andamios nHAC / PLGA híbridos con óxido de grafeno facilitan la proliferación de células MC3T3-E1
- Estudio comparativo de las propiedades electroquímicas, biomédicas y térmicas de nanomateriales naturales y sintéticos
- Reduzca la sensibilidad de CL-20 mejorando la conductividad térmica a través de nanomateriales de carbono
- Ajuste del modo de movimiento deslizante de los nanotubos de carbono mediante grupos de hidroxilo