


Asimilación y acción de nano-Se en aves de corral y otros animales monogástricos:¿Es la microbiota intestinal una respuesta?
Resumen
Recientemente, se ha publicado en Nanoscale Research Letters un artículo de revisión completo dedicado a las funciones del nano-Se en la nutrición del ganado y los peces. Los autores describieron con gran detalle un tema relacionado con la producción de nano-Se y sus posibles aplicaciones en la industria animal y la medicina. Sin embargo, no se describieron los mecanismos moleculares de la acción de nano-Se y la cuestión de cómo se convierte nano-Se en selenoproteínas activas no está resuelta. Parece probable que la microbiota intestinal pueda convertir nano-Se en selenita, H 2 Se o Se-fosfato con la siguiente síntesis de selenoproteínas. Esta posibilidad debe estudiarse más en detalle, y las ventajas y desventajas del nano-Se como fuente de Se en la nutrición de animales / aves / peces aguardan evaluaciones críticas.
Antecedentes
Recientemente, se ha publicado en Nanoscale Research Letters [1] un artículo de revisión exhaustivo dedicado a las funciones del nano-Se en la nutrición del ganado y los peces. Los autores describieron con gran detalle un tema relacionado con la producción de nano-Se y sus posibles aplicaciones en la industria animal y la medicina. De hecho, está bien establecido que muchas moléculas presentadas como nanopartículas tienen un comportamiento inusual debido a las nuevas propiedades de tales partículas. De hecho, en la mayoría de los trabajos publicados relacionados con la nutrición animal, las nanopartículas tienen un tamaño inferior a 100 nm. En tal estado, el tamaño ultrapequeño de las partículas les permite atravesar muchas barreras biológicas y ser utilizadas como un sistema de liberación de varios elementos, incluidos Ag, titanio, Se y otros.
En este sentido, el Se es especialmente interesante, ya que está comprobado que la mayoría de áreas a nivel mundial son deficientes en este elemento [2]. La deficiencia en muchos casos es una creación humana, ya que el contenido de Se en los suelos varía mucho y, lo que es más importante, la disponibilidad de Se de los suelos es aún más variable. De hecho, la acidificación del suelo como resultado de las prácticas agrícolas, así como el uso de fertilizantes que contienen azufre, disminuyó sustancialmente la disponibilidad de Se para las plantas y conduce a un bajo consumo de Se por parte del ganado, donde los granos (trigo, maíz, cebada) y semillas oleaginosas (soja) constituyen una parte importante de la dieta. Por lo tanto, la suplementación con Se de las dietas comerciales de aves de corral, animales de granja y pescado se convirtió en una práctica común y se utiliza desde la década de 1970. De hecho, las premezclas de vitaminas y minerales son una fuente importante de Se para la producción comercial de huevos, carne / pescado y leche.
Lo que es relevante para la historia de nano-Se es la fuente de Se en los suplementos. De hecho, la principal forma suplementaria de Se es el selenito de sodio, que es un subproducto de la producción de cobre. Con más de 40 años de uso, el selenito de sodio ha demostrado claramente sus ventajas y desventajas. De hecho, debido a la suplementación comercial de Se para piensos, la deficiencia de Se en el ganado con signos clínicos de deficiencia prácticamente desapareció. La excepción son los rumiantes, donde el selenito de sodio puede precipitar en un ambiente ruminal ácido, así como debido a limitaciones técnicas del uso de suplementos alimenticios; La deficiencia de se todavía se observa en todo el mundo y se utilizan aplicaciones como inyecciones de selenio y bolos. Sin embargo, como se menciona correctamente en la revisión antes mencionada, el Se es extremadamente importante en condiciones de estrés comercialmente relevantes, especialmente para la genética moderna de animales de granja y aves de corral altamente productivos.
Hoy en día, la industria animal está pasando de la prevención de las deficiencias de nutrientes a satisfacer los requisitos exactos de los animales en nutrientes importantes, incluido el Se. De hecho, "nutrición precisa" es un término que describe este concepto. En tales condiciones, se ha hecho evidente que el selenito de sodio (así como el selenato) tiene una serie de desventajas como complemento alimenticio. En primer lugar, es un compuesto bastante reactivo y se puede reducir en el pienso / premezcla a una forma metálica no disponible mediante varios nutrientes, incluido el ácido ascórbico y algunos ingredientes del pienso. También puede disolverse en la humedad del alimento y convertirse en compuestos volátiles que se perderán. En segundo lugar, el selenito de sodio posee propiedades prooxidantes que dependen de la dosis, lo que podría tener un efecto negativo en el intestino del animal / pollo. Finalmente, el selenio en forma de selenito de sodio se transfiere pobremente a los huevos, a través de la placenta a los fetos, que no es capaz de acumular reservas de Se en el cuerpo que pueden usarse en condiciones de estrés cuando aumenta el requerimiento de Se pero el consumo de alimento generalmente disminuye. El selenito de sodio también es tóxico para animales y humanos en exceso. Sin embargo, no estamos de acuerdo con la conclusión de la revisión mencionada [1] de que la toxicidad por Se es un problema para la industria animal. Solo está relacionado con un error humano en el cálculo o en la mezcla de alimentos. La ventana segura para el selenio es bastante estrecha (la dosis habitual de suplementos de Se para aves y cerdos es de aproximadamente 0,3 ppm, mientras que se observaría un efecto negativo con dosis de selenito de sodio superiores a 1 a 2 ppm), pero los equipos modernos de fábricas de piensos ofrecen una oportunidad de una buena mezcla de alimentos evitando problemas de toxicidad.
Los avances en las ciencias analíticas fueron una fuerza impulsora en el descubrimiento de que la forma principal de Se en los principales ingredientes de los piensos es SeMet, que comprende más del 50% del Se total en maíz, soja, trigo, cebada, etc. [2]. Por lo tanto, durante la evolución, el sistema digestivo de los animales se adaptó a esta forma de selenio y, como resultado, SeMet se asimila de manera más eficiente a partir de la dieta y la incorporación no específica a las proteínas corporales genera reservas de Se. De hecho, apareció en el mercado una gama de fuentes de Se orgánico que incluyen Se-Yeast, SeMet puro y OH-SeMet (ácido 2-hidroxi-4- (metiltio) butanoico-Se), con OH-SeMet para mostrar la máxima eficacia.
El principal desconocimiento en el tema nano-Se está relacionado con su metabolismo y particularmente su conversión a H 2 Se con la siguiente síntesis e incorporación de SeCys en selenoproteínas. En la revisión [1], solo se mencionan muy pocas selenoproteínas, incluida la glutatión peroxidasa, mientras que al menos se identifican 25 selenoproteínas en humanos y animales. Se acepta generalmente que el papel principal del Se en la nutrición humana / animal está relacionado con la síntesis de selenoproteínas que poseen propiedades catalíticas únicas y más de la mitad de ellas están involucradas en el mantenimiento del equilibrio redox y las defensas antioxidantes [3]. En la revisión antes mencionada [1], las propiedades antioxidantes directas (reducción de ROS) del nano-Se se mencionan como un posible mecanismo de su acción. Sin embargo, la concentración de Se en los principales tejidos de animales / aves de corral, incluidos el hígado y los músculos, por lo general no supera los 800-900 ng / g de tejido fresco, que está en el rango de 10 μM, y en el plasma, el nivel de Se es de aproximadamente 0.2-0.3 μg / ml o 2-3 μM, mientras que en el trabajo más citado dedicado a las propiedades antioxidantes de nano-Se in vitro, las concentraciones de Se fueron probadas y mostraron efectos antioxidantes 5-10 veces más altos [4]. Además, para que un compuesto antioxidante sea un eliminador de radicales libres eficaz, es importante tener la concentración correcta del antioxidante en el lugar correcto en el momento adecuado. Esto complicaría aún más el problema y, por lo tanto, parece poco probable que nano-Se pueda tener un efecto AO directo en los sistemas biológicos.
Por lo tanto, al igual que otras formas de selenio utilizadas en dietas animales, un efecto antioxidante de nano-Se está relacionado con la expresión génica de selenoproteína y la síntesis de proteínas. De hecho, en la revisión [1], hay varias referencias que confirman un efecto positivo de nano-Se sobre la actividad GSH-Px. Ahora la pregunta es cómo nano-Se se convierte en selenoproteínas activas. En la revisión [1], hay una sugerencia (sin una referencia) de que nano-Se puede convertirse en selenofosfato con la siguiente síntesis de proteína Se. Esta sugerencia debe probarse experimentalmente. La segunda sugerencia de que nano-Se puede convertirse en SeMet es fundamentalmente errónea, ya que SeMet no se puede sintetizar en el cuerpo humano / animal; solo las plantas y las bacterias pueden producirlo [5].
Hay sugerencias muy atractivas de que la microbiota intestinal puede oxidar nano-Se en selenito / selenato o reducirlo a H 2 Se con la siguiente síntesis de selenoproteínas [6, 7]. Recientemente, se han proporcionado algunas pruebas experimentales para demostrar que las partículas de nano-Se pueden disolverse y oxidarse a oxoaniones inorgánicos de Se en el intestino en presencia de microbiota antes de su absorción [6]. Además, se propuso un diagrama esquemático hipotético que muestra un ciclo dinámico intracelular de nanopartículas de selenio endógeno (SeNP) [7]. De hecho, se sugirió que los radicales superóxido podrían volver a oxidar el selenio elemental en selenito. También existe la posibilidad de que la microbiota intestinal pueda realizar / acelerar este proceso. De hecho, se ha demostrado que ocurren en la naturaleza cuatro transformaciones biológicas principales del Se, que incluyen la reducción, oxidación, metilación y desmetilación [8]. Los datos acumulados durante las últimas tres décadas indican claramente que los microorganismos desempeñan un papel importante en el ciclo del selenio en el medio ambiente al participar tanto en reacciones de oxidación como de reducción [9]. Curiosamente, la oxidación microbiana de Se 0 a Se 4+ por un grupo de bacterias autótrofas no identificadas fue descubierto hace más de 90 años [10]. Además, la oxidación del selenio elemental a selenito por una bacteria heterotrófica, Bacillus megaterium , aislado del suelo se informó mucho más tarde [11]. De hecho, Se 0 Se demostró que la oxidación en los suelos ocurre a un ritmo relativamente lento y que es en gran parte de naturaleza biótica y produce tanto SeO 3 2− y SeO 4 2− [12]. Además, la oxidación microbiana del selenio elemental (Se 0 ) por quimioheterótrofos y tiobacilos quimioautótrofos se confirmó mediante el uso de 75 Se 0 como trazador [13]. Los autores demostraron que las lechadas del suelo podían oxidar Se 0 con SeO 3 2− y SeO 4 2− formación. Curiosamente, la inactivación microbiana en el suelo mediante autoclave o tratamientos químicos inhibió el proceso. Además, los cultivos de bacterias oxidantes de azufre Thiobacillus Se ha demostrado que ASN-1 realiza la oxidación de Se (0) con enzimas que se utilizan para generar energía a partir de compuestos reducidos de azufre [13]. Además, la bacteria heterotrófica que puede oxidar Mn (II) o Fe (II) ( Leptothrix Se demostró que MnB1) oxida Se 0 con la formación de SeO 3 2− como el principal producto de reacción. Curiosamente, se demostró que la reacción depende de un donante de electrones como el acetato o la glucosa [13]. La oxidación de Se 0 por diversas bacterias no se ha abordado por completo y es un área de importantes oportunidades de investigación. De hecho, una gran variedad de microbios que residen en el intestino proporciona las condiciones necesarias para varias conversiones de Se. Por ejemplo, cuando las partículas de nano-Se se incubaron con bacterias del ácido láctico ( Lactobacillus delbrueckii subsp . bulgaricus LB-12), se produjeron compuestos orgánicos de Se (principalmente SeCys y SeMet) y también las nanopartículas de Se se disolvieron parcialmente y se transformaron de forma no metabólica en selenio inorgánico, probablemente con la ayuda de sustancias excretadas por la pared celular de las bacterias [14].
Alternativamente, el selenio elemental se puede reducir / convertir en seleniuro por una bacteria que respira selenito (por ejemplo, Bacillus selenitireducens ). La reacción antes mencionada llevada a cabo por las bacterias con la oxidación incompleta del lactato donante de electrones a acetato de la siguiente manera se presentó de la siguiente manera [7]:
$$ {\ mathrm {C}} _ 2 {\ mathrm {H}} _ 4 {\ mathrm {OHCOO}} ^ {-} + 2 {\ mathrm {Se}} ^ 0 + 2 {\ mathrm {H}} _ 2 \ mathrm {O} \ to {\ mathrm {C} \ mathrm {H}} _ 3 {\ mathrm {C} \ mathrm {OO}} ^ {-} + 2 {\ mathrm {H} \ mathrm {Se}} ^ {-} + {{\ mathrm {H} \ mathrm {CO}} _ 3} ^ {-} + 3 {\ mathrm {H}} ^ {+} $$Por tanto, las energías libres para la reacción (∆G I ) se muestra como - 2.8 kcal / mol e - . Esto muestra que en Bacillus selenitireducens , el mecanismo de reducción implica la conservación de energía mediante el uso de enzimas disimilatorias específicas de Se [15]. De hecho, una bacteria que respira selenita, Bacillus selenitireducens , podría producir niveles significativos de Se - 2 (como HSe- acuoso) usando Se 0 como sustrato [16]. Anteriormente se demostró que el selenio rojo era reducido por el acidófilo obligado Thiobacillus ferrooxidans en condiciones ácidas (pH 3), anaeróbicas con H 2 Producción de Se a una tasa de 0,03 µmol / mg de proteína / h [17]. Además, otra bacteria anaeróbica Veillonella atypica demostró ser capaz de reducir los oxianiones de selenio para formar selenio elemental con su reducción adicional por parte de la bacteria para formar seleniuro reactivo [18]. Curiosamente, más de 45 años antes, se describió que los extractos de Micrococcus lactilyticus ( Veillonella alcalescens ) pudieron reducir cuantitativamente el selenio coloidal en H 2 Se [19]. Recientemente, los genes que codifican las proteínas YedE y YedF se han considerado como nuevos genes candidatos implicados en el metabolismo del Se en procariotas, incluidas las bacterias [20]. De hecho, tanto YedE, un transportador de Se predicho, como YedF, una proteína redox, podrían participar en la transformación metabólica del selenio en las células bacterianas. Por lo tanto, se comprobó la capacidad principal de varios microbios para oxidar o reducir el selenio elemental (Cuadro 1); sin embargo, es necesario realizar más investigaciones para responder a la pregunta de si tales reacciones tienen lugar en el intestino del animal. Curiosamente, de todas las especies bacterianas mencionadas en la Tabla 1, las bacterias anaerobias gramnegativas del género Veillonella merecen una atención especial . De hecho, Veillonellae se encuentran en el tubo digestivo de animales de sangre caliente [21]. De hecho, en los animales destinados a la alimentación, Veillonella Las cepas se detectan regularmente como habitantes autóctonos de todas las secciones del tracto gastrointestinal ([22] y las referencias allí), incluido el tracto gastrointestinal superior [23] y el ciego [24] de los pollos. De hecho, se necesita una investigación adicional para dilucidar el destino y los mecanismos de las posibles conversiones de nano-Se en el tracto gastrointestinal de los animales. El metabolismo de nano-Se depende de la composición de las nanopartículas, incluido el agente de recubrimiento de nanopartículas. En la figura 1 se muestra un esquema propuesto de participación de nano-Se en la síntesis de selenoproteínas, y una comprensión básica del metabolismo de nano-Se, incluida la absorción, distribución y eliminación, es de gran importancia en las ciencias animales / avícolas [25].
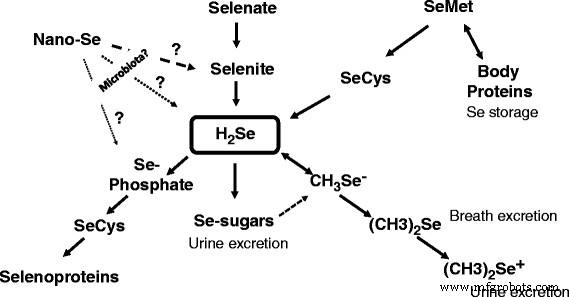
Modelo esquemático que muestra conversiones metabólicas de diversas formas de Se en animales. Se sugiere que la microbiota intestinal podría convertir nano-Se en selenita, H 2 Se o Se-fosfato con la siguiente síntesis de SeCys e incorporación en selenoproteínas
El tracto gastrointestinal de mamíferos / aves alberga billones de microorganismos comensales, conocidos colectivamente como microbiota [26]. Por ejemplo, el tracto intestinal del pollo se compone de duodeno, yeyuno, íleon, ciego y colon, y existen diferencias significativas en la concentración y composición de la microbiota entre las secciones del intestino antes mencionadas [27]. Curiosamente, el ciego se caracteriza por la comunidad microbiana más compleja dominada por los phyla Firmicutes , Bacteroidetes , Actinobacterias y proteobacterias [28]. Por otro lado, a nivel de género, se demostró que los principales géneros microbianos en todas las secciones intestinales son Lactobacillus , Enterococcus , Bacteroides y Corynebacterium [27]. Además, Bacteroides demostró ser el grupo dominante en el ciego, mientras que Lactobacillus fue predominante en las secciones del intestino delgado (duodeno, yeyuno e íleon; [27]). En este complejo ecosistema intestinal, podría haber una variedad de microbios capaces de facilitar la oxidación / reducción de nanopartículas de Se, y esta suposición espera una mayor investigación.
Por un lado, algunos microbios usarían Se para sus propias necesidades para sintetizar selenoproteínas microbianas y competir con el huésped por este elemento. Es bien sabido que el Se es un elemento importante para una variedad de organismos en casi todos los filos bacterianos; sin embargo, parece probable que solo un tercio de las bacterias caracterizadas utilicen este elemento en su metabolismo [29]. De hecho, en la deficiencia de Se, existe una competencia entre las bacterias y el huésped por el selenio disponible, y los animales libres de gérmenes tienen un requerimiento de selenio menor que los animales colonizados convencionalmente [30]. Resultados recientes indican que la suplementación dietética con Se puede afectar tanto la composición como la diversidad de la microbiota existente y el establecimiento de la microflora gastrointestinal [31]. Por ejemplo, se demostró que los terneros destetados alimentados con heno de alfalfa enriquecido con selenio tienen una microbiota nasal enriquecida en comparación con los animales de control [32]. Los posibles efectos del Se en diversas formas y concentraciones sobre la microbiota intestinal aguardan una mayor investigación.
Por otro lado, parece probable que haya una absorción activa de Se por parte de la microbiota, y este proceso depende de la forma de Se utilizada. De hecho, se demostró que la captación de SeMet por la microbiota del colon es mucho más eficaz en comparación con la captación de selenato [33]. Sin embargo, no se ha investigado si la composición y concentración de bacterias afectan la absorción de compuestos de selenio en el intestino. En general, se acepta que la microbiota intestinal es responsable de la excreción del selenio en exceso mediante su metilación y formación de Se elemental [34]. Curiosamente, en ratas alimentadas con SeMet, esta forma de Se se encontró en todos los segmentos del intestino. Sin embargo, el nivel de Se en el íleon, ciego y colon fue significativamente más alto que en las secciones correspondientes de ratas después del tratamiento con probióticos [34]. Esto podría significar que Streptococcus salivarius , Lactobacillus rhamnosus , Lactobacillus acidophilus y Bifidobacterium lactis se administraron al intestino con metabolismo de Se afectado por probióticos principalmente en el intestino distante. De hecho, se debe identificar el efecto de diferentes bacterias en la conversión de Se para comprender las funciones de cada segmento del intestino en el metabolismo y la asimilación del Se. Además, el reflujo del líquido de la parte superior del intestino grueso al intestino delgado puede ser responsable de la absorción de H 2 Se y otras formas de Se del intestino. De hecho, recientemente se ha demostrado que los pollos se caracterizan por contracciones peristálticas inversas que podrían mover el marcador de la cloaca a la molleja [35].
Se podría esperar una participación directa de nano-Se en la síntesis de selenoproteínas, ya que en cultivos celulares nano-Se aumentaron las selenoproteínas (actividad GSH-Px y TR). Sin embargo, la comprensión reciente de la prioridad de expresión de selenoproteínas podría complicar este problema. De hecho, muchas selenoproteínas están reguladas por estrés oxidativo. En particular, se demostró que GSH-Px1, GSH-Px4 y TR1 estaban regulados al alza en respuesta al estrés oxidativo [36], y dicha respuesta era más pronunciada cuando el suministro de Se era limitante. Cabe mencionar que la respuesta antes mencionada depende también del nivel de estrés oxidativo, porque es cierto en el estrés oxidativo leve, pero a un nivel extremadamente alto, se activan algunos otros mecanismos [2].
Bien podría ser que en cultivo celular, el aumento de expresión / actividades de tales selenoproteínas sea una respuesta a un estrés oxidativo creado por nano-Se, pero no un reflejo de un suministro mejorado de Se. Por lo tanto, se debe tener precaución al interpretar los resultados basados en estudios de cultivos celulares. Una confirmación adicional de los cambios relacionados con el estrés en los sistemas biológicos debido a la suplementación con nano-Se provino de un estudio reciente que muestra que el nano-Se biogénico podría activar el factor nuclear (derivado del eritroide-2) -como 2 (Nrf2) y aumentar la expresión de sus genes posteriores, responsables de la síntesis de antioxidantes en formas dependientes de la dosis y del tiempo [37]. Además, los autores encontraron que la eliminación de Nrf2 bloqueaba significativamente el efecto antioxidante de tales nanopartículas de Se.
Al analizar críticamente los efectos positivos de la suplementación con nano-Se sobre el desempeño productivo y reproductivo de aves de corral, animales de granja y peces, es necesario tener en cuenta que en la mayoría de los casos, el Se no mejoró el desempeño sino que previno el deterioro del desempeño debido a factores ambientales o limitaciones nutricionales. En muchos casos, los experimentos con animales se llevaron a cabo con un nivel de fondo de Se extremadamente bajo y, por lo tanto, agregar Se en cualquier forma disponible puede tener efectos positivos.
Los desafíos de la comercialización de nano-Se para su uso como aditivo alimentario podrían ser los siguientes:
- 1.
Es necesario comprender los mecanismos moleculares de absorción, asimilación y acción de nano-Se a nivel celular, subcelular y genético. Sin tales datos, sería difícil tener resultados reproducibles y encontrar explicaciones adecuadas de los efectos observados. Por ejemplo, en un artículo publicado recientemente, solo unos pocos genes (18 proteínas y ninguna de ellas está directamente relacionada con el metabolismo del Se) se vieron afectados en el hígado debido a una sobredosis de nano-Se en pollos [38], mientras que se sabe que otros formas de Se dietético, incluido SeMet, pueden afectar a unos cientos de genes.
- 2.
Es importante comprender si nano-Se puede generar reservas de Se en el cuerpo, como SeMet, y si esas reservas están disponibles en condiciones de estrés.
- 3.
Es necesario diseñar una tecnología capaz de proporcionar partículas de nano-Se del mismo tamaño, estabilidad y capacidad de almacenamiento razonablemente buena (al menos 6-12 meses) [2].
- 4.
Es probable que los probióticos microbianos puedan ser útiles en la conversión de nano-selenio usado como aditivo alimentario. Por un lado, estos probióticos pueden contener microorganismos específicos que ayudan a la asimilación de nano-Se en el intestino (p. Ej., Veillonella especies). Por otro lado, los probióticos enriquecidos con Se podrían ser otra posibilidad [34, 39]
- 5.
Los efectos secundarios, los riesgos y las preocupaciones ambientales deben abordarse en su totalidad. De hecho, se requieren más investigaciones para informar a los responsables políticos y los organismos reguladores sobre el potencial nanotoxicológico de nano-Se [40]. En particular, un tamaño muy pequeño de partículas en forma seca hace que el producto sea muy polvoriento, y el comportamiento inusual de las partículas de nano-Se, una vez en el cuerpo, da una advertencia sobre la principal preocupación por la protección de los trabajadores de las fábricas de piensos. Dado que la microbiota intestinal es responsable de la excreción del selenio en exceso por su metilación y formación de Se elemental [34], la mejora / modulación de la microbiota podría abrir un nuevo horizonte para hacer frente a la posible toxicidad del nano-Se.
- 6.
Los efectos positivos de nano-Se en la nutrición animal no deberían eclipsar las posibles consecuencias perjudiciales de su uso. De hecho, el comportamiento de las nanopartículas en diversas condiciones podría diferir sustancialmente, y antes de que entendamos cómo controlar ese comportamiento, el uso de nano-Se a gran escala industrial no debería ser posible. Por ejemplo, al considerar la absorción de nano-Se, es necesario mencionar el llamado efecto caballo de Troya, cuando las nanopartículas pueden tener propiedades que mejoran la permeación de otras sustancias en el intestino [41]. Esto podría crear algunos problemas, ya que existe una variedad de compuestos "no deseados" en el alimento y el intestino protege contra su absorción. De hecho, el comportamiento de nano-Se en el intestino merece una mayor investigación.
- 7.
En la revisión antes mencionada [1], se presta una atención sustancial a las propiedades antimicrobianas y anticancerígenas del nano-Se, y parece probable que las propiedades únicas de las nanopartículas puedan ayudar a combatir diversas enfermedades. De hecho, nano-Se puede considerarse como un nuevo fármaco para su uso en diversas afecciones médicas, incluida la terapia contra el cáncer, mientras que su uso como aditivo alimentario eficaz es bastante cuestionable. La investigación futura debe responder a esas preguntas e inquietudes importantes.
Conclusión
Durante los últimos años, se publicaron una serie de artículos dedicados a nano-Se y el tema se está desarrollando rápidamente. Sin embargo, antes de que esta forma de Se pueda encontrar una forma de producción comercial de aves / animales, es necesario comprender y explicar cómo el nano-Se se convierte en selenoproteínas activas. Uno de los posibles mecanismos / vías de acción de nano-Se podría estar mediado por la microbiota intestinal que podría convertir nano-Se en selenita, H 2 Se o Se-fosfato con la siguiente síntesis de selenoproteínas (Fig. 1). Existe alguna evidencia de que en la naturaleza, las bacterias podrían reducir u oxidar el Se metálico (Se 0 ) con la producción de Se −2 o Se +4 y Se +6 respectivamente. Entre los microorganismos implicados en los cambios redox de Se, el género Veillonella merece una atención especial debido a la presencia de tales bacterias en el intestino de los animales destinados a la alimentación, incluido el pollo. La posibilidad de la participación de la microbiota intestinal en la asimilación y el metabolismo de nano-Se debe investigarse más en detalle, y las ventajas y desventajas de nano-Se como fuente de Se en la nutrición de animales / aves de corral esperan más evaluaciones críticas.
Nanomateriales
- 3 razones por las que DevOps y la nube se necesitan mutuamente; DevCloudOps
- Tornillos de PVDF, pernos hexagonales, tuercas hexagonales, otros sujetadores y piezas personalizadas
- Las plantas y animales obtenidos por reproducción biológica ya no son patentables en la OEP
- 3 Ventajas de la simulación de robots para fabricantes y otros usuarios de robots
- Automatización de procesos en SAP S/4HANA®:esperanzas, impulsores y acción
- Diferencias de Nexus Integra con otras plataformas IoT y Big Data
- ¿Qué software y otras herramientas utilizan los ingenieros de PCB?
- Composición de níquel 718 y otras aleaciones clave
- Las ventajas de usar restos de metal de aluminio, gotas y otros cortos de metal
- Principales preocupaciones de seguridad en camiones y cómo evitarlas
- Centros de mecanizado horizontal (HMC):precio, costos y otras consideraciones