


ARN largo no codificante MALAT1 / microARN-143 / VEGFA El eje de señal modula el aneurisma intracraneal inducido por lesión del endotelio vascular
Resumen
Las funciones de algunos ARN no codificantes largos (lncRNA) en el aneurisma intracraneal (IA) se han investigado en muchos estudios. El objetivo de este estudio es dilucidar el mecanismo del transcrito 1 de adenocarcinoma de pulmón asociado a metástasis de lncRNA (MALAT1) / microRNA-143 (miR-143) / factor de crecimiento endotelial vascular A (VEGFA) en el eje de la señal en la IA inducida por lesión endotelial vascular . Se detectó la expresión de MALAT1, miR-143 y VEGFA en tejidos IA y tejidos arteriales normales. Se detectaron metaloproteinasa de matriz 9 (MMP-9) en tejidos, factor de von Willebrand (vWF) en suero y tejidos, y endotelina-1 (ET-1) en suero. Las ratas IA modeladas se inyectaron con MALAT1 silenciado o sobreexpresado para detectar lesión endotelial vascular. Se extrajeron y transfectaron células endoteliales vasculares de pacientes con IA con MALAT1 silenciado o sobreexpresado para verificar los impactos de MALAT1 en la viabilidad celular y la apoptosis. Las conexiones entre MALAT1, miR-143 y VEGFA se verificaron mediante predicción en línea, actividad de luciferasa y ensayos de extracción de ARN. Se encontró sobreexpresión de MALAT1 y VEGFA y mala expresión de miR-143 en tejidos IA. La regulación a la baja de MALAT1 inhibió la presión arterial, la expresión de ET-1, vWF y MMP-9, así como el índice apoptótico de las células endoteliales vasculares de ratas con IA. MALAT1 regulado a la baja inhibió la apoptosis y promovió la viabilidad de las células endoteliales vasculares en IA. MALAT1 se unió a VEGFA dirigido a miR-143 y miR-143. Este estudio sugiere que MALAT1 eleva la expresión de VEGFA a través de la unión competitiva a miR-143, lo que aumenta la apoptosis y atenúa la viabilidad de las células endoteliales vasculares en IA.
Introducción
El aneurisma intracraneal (IA), también conocido como aneurisma cerebral, es causado por el aumento de la presión intracraneal inducida por el ensanchamiento anormal local de la cavidad de la arteria cerebral o la pared arterial [1]. La AI es una enfermedad grave con una alta mortalidad y morbilidad, y la tasa de prevalencia es de alrededor del 1-3% en la población general [2]. Las principales características clínicas de la AI son el vasoespasmo cerebral, la hemorragia cerebral espontánea y la parálisis del nervio motor ocular común [3]. Hasta ahora, los factores de riesgo comunes que conducen a la aparición y desarrollo de AI incluyen trastornos hemodinámicos, genes, envejecimiento, infecciones y factores congénitos [4]. Los principales tratamientos clínicos, principalmente el clipaje quirúrgico y / o el enrollamiento endovascular, sirven para prevenir la rotura del aneurisma [5]. Sin embargo, el mecanismo detallado que subyace a la EI aún está por dilucidarse, lo que refleja la necesidad urgente de métodos más efectivos en la gestión de la EI.
Los ARN largos no codificantes (lncRNA) tienen más de 200 nucleótidos que pertenecen a una especie de ARN no codificantes [6]. Se ha informado de que la expresión del lncRNA 1 asociado a la detención del crecimiento de lncRNA en IA está regulada negativamente y el transcrito 1 de adenocarcinoma de hongos (MALAT1) es un lncRNA altamente enriquecido y ampliamente expresado cuya longitud es de aproximadamente 8000 nt [7]. Se ha documentado que MALAT1 modula la disfunción del músculo liso en el aneurisma de la aorta torácica [8]. Además, un estudio ha presentado la expresión anormal de lncRNA y ARN mensajero (mRNA) en IA, y las redes de coexpresión de lncRNA-mRNA proporcionan pistas para encontrar la patogénesis de IA [9]. Se ha sugerido que MALAT1 promueve la expresión de osterix y modula la diferenciación osteogénica mediante la focalización en la expresión de miARN-143 (miR-143) en células madre mesenquimales de la médula ósea humana [10]. Un estudio también ha presentado el papel del grupo miR-143/145 en revertir la regulación del factor 5 similar a krüppel en las células del músculo liso y su contractilidad y proliferación en AI [11]. Según Feng et al., La regulación a la baja de miR-143/145 y un nivel más alto de metaloproteinasa de matriz-9 (MMP-9) durante la circulación puede estar relacionado con la formación y ruptura de AI [12]. Un análisis ha revelado que los miARN más incontrolados (miR-143 y miR-145) son comunes a los genes diana que son vías de señal, como el factor de crecimiento endotelial vascular (VEGF), y otros genes que regulan el movimiento o la adhesión celular [13]. . Un estudio ha revelado la importancia predictiva de las variaciones del factor de crecimiento endotelial vascular A (VEGFA) en la AI [14]. Por lo tanto, buscamos evaluar el mecanismo del eje de señal MALAT1 / miR-143 / VEGFA en IA inducido por lesión endotelial vascular.
Materiales y métodos
Declaración de ética
El estudio fue respaldado por la Junta de Revisión Institucional del Primer Hospital Afiliado de la USTC, División de Ciencias de la Vida y Medicina de la Universidad de Ciencia y Tecnología de China y siguió los principios de la Declaración de Helsinki. Los participantes proporcionaron su consentimiento informado por escrito en este estudio. Todos los experimentos con animales se ajustaron a la Guía para el cuidado y uso de animales de laboratorio de los Institutos Nacionales de Salud. El protocolo fue autorizado por el Comité de Ética de Experimentos con Animales del Primer Hospital Afiliado de la USTC, División de Ciencias de la Vida y Medicina de la Universidad de Ciencia y Tecnología de China.
Temas de estudio
Se seleccionaron para nuestros experimentos veinte pacientes IA (grupo IA) que habían sido diagnosticados mediante un examen de imágenes y habían recibido un clip neuroquirúrgico en el Primer Hospital Afiliado de la USTC, División de Ciencias de la Vida y Medicina, Universidad de Ciencia y Tecnología de China. Había 11 hombres y 9 mujeres que tenían 43.27 ± 6.25 años. Se tomaron muestras de los tejidos IA. Mientras tanto, los tejidos vasculares arteriales corticales temporales del polo temporal se resecaron de 20 pacientes (grupo de control) con epilepsia del lóbulo temporal causada por esclerosis de la amígdala y el hipocampo. Los tejidos resecados se examinaron como tejidos arteriales normales por histopatología después de la operación, y había 13 hombres y 7 mujeres de 44,18 ± 5,91 años. No se reconoció ninguna discrepancia significativa en el género y la edad entre el grupo IA y el grupo de control (tanto P > 0,05). Se obtuvieron muestras de sangre venosa (2 tubos) de todos los sujetos en ayunas a la misma hora la mañana antes de la operación.
Establecimiento de modelos de rata de IA
Se criaron sesenta ratas macho Sprague-Dawley de grado limpio (Hunan SJA Laboratory Animal Co., Ltd., Hunan, China), con un peso de entre 200 y 250 g, durante 7 días (25 ± 2 ° C, humedad relativa de 65-70 %, 12 h de ciclo de luz y oscuridad, agua libre y ingesta alimentaria). Las ratas se dejaron caer con elastasa pancreática porcina a la arteria carótida externa y alrededor de la pared de la arteria de bifurcación. La arteria carótida externa se ligó con dos líneas quirúrgicas en la rama de la arteria carótida externa de aproximadamente 1,5 mm. La arteria carótida externa se cortó entre las dos líneas para formar un aneurisma de la carótida interna en el segmento ciego de la arteria carótida externa. Las ratas se alimentaron con solución salina al 1% durante 1 semana después de la operación. Se realizó angiografía cerebral al cabo de 1 mes y se observó formación de aneurisma.
Después del establecimiento de modelos de ratas IA, se distribuyeron al azar 50 ratas en un grupo en blanco ( n =10, las ratas modeladas se trataron con inyección estereotáctica de 100 μL de solución salina tamponada con fosfato (PBS) una vez al día), grupo de control negativo (NC) de ARN de horquilla corta (sh) ( n =10, las ratas modeladas se trataron con una inyección estereotáctica de 100 μL de sh-MALAT1 NC una vez al día), grupo sh-MALAT1 ( n =10, las ratas modeladas se trataron con inyección estereotáctica de 100 μL de plásmido sh-MALAT1 una vez al día), grupo de sobreexpresión (Oe) -NC ( n =10, las ratas modeladas se trataron con inyección estereotáctica de 100 μL de plásmido Oe-MALAT1 NC una vez al día) y el grupo Oe-MALAT1 ( n =10, las ratas modeladas se trataron con inyección estereotáctica de 100 μL de plásmido Oe-MALAT1 una vez al día) [15]. Los plásmidos anteriores fueron compuestos por Shanghai Genechem Co., Ltd. (Shanghai, China).
Prueba de presión arterial de ratas
La presión arterial de la arteria de la cola de las ratas se midió en la 1ª, 4ª y 12ª semana después de la operación. Antes de medir la presión arterial, las ratas se colocaron en un dispositivo de calentamiento de temperatura constante durante un momento para evitar la alteración de la temperatura externa. En segundo lugar, las ratas se mantuvieron en silencio durante varios minutos en una jaula especial para ratas para evitar la interferencia de la actividad. Si la presión arterial variaba en gran medida, se determinaba dos o tres veces en diferentes momentos para obtener el valor promedio.
Adquisición de tejido de aneurisma
Después de 3 meses, las ratas se anestesiaron con pentobarbital sódico al 1% (40 mg / kg) mediante inyección intraperitoneal para obtener muestras de sangre de las venas. Se sacrificó a las ratas y se abrió el tórax, se intubó el ventrículo izquierdo en la aorta y se cortó la cava para liberar la sangre. Mientras tanto, se utilizaron 30 ml de solución salina normal que contenía heparina sódica para una perfusión cardíaca rápida para enjuagar la sangre, y luego se inyectaron en el cerebro 10 ml de poliformaldehído al 10% / PBS 0,1 M (pH 7,4). Después de la perfusión y la fijación, se abrió el cerebro de la rata. Se observó cuidadosamente la circulación arterial en la base del cráneo, se separó el tejido del aneurisma y se observaron los cambios del aneurisma al microscopio.
Ensayo inmunoabsorbente ligado a enzimas (ELISA)
Los índices relacionados con el suero se probaron mediante el kit ELISA. Las muestras de sangre recolectadas se colocaron en un termostato a 37 ° C durante 1 hy se centrifugaron a 3000 r / min durante 10 min. La detección de la expresión de endotelina-1 (ET-1) y factor von Willebrand (vWF) se llevó a cabo de acuerdo con las instrucciones del kit (todos los kits se compraron en el Instituto de Bioingeniería NanJing JianCheng, Jiangsu, China).
Tinción con hematoxilina-eosina (HE)
Las muestras se fijaron con formalina al 10% durante más de 24 hy se conservaron en bloques de parafina. Los bloques de parafina se desparafinaron con xileno durante 20 min, se deshidrataron con una serie descendente de alcohol en gradiente (100%, 95%, 80%, 75%) durante 1 min y se tiñeron con hematoxilina durante 10 min. Luego, los tejidos se enjuagaron con agua destilada, se diferenciaron con ácido clorhídrico etanol durante 30 sy se remojaron en agua tibia a 50 ° C durante 5 min. Teñidos con solución de eosina, los tejidos se enjuagaron con agua destilada, se deshidrataron con alcohol al 70% y 90%, se aclararon con xileno y se sellaron con goma neutra. La morfología de los tejidos se observó bajo un microscopio de alta potencia.
Observación con microscopio electrónico de transmisión
Los tejidos sobrantes se fijaron con dialdehído glutárico al 2,5% y ácido osmítico al 1%, se deshidrataron y luego se embebieron con resina Epon812. Las secciones semifinas se tiñeron con azul de tolueno, se recortaron y se transformaron en secciones ultrafinas. Las secciones se tiñeron con acetato de uranilo y citrato de plomo y se observaron con un microscopio electrónico de transmisión JME-2000EX (Hitachi High-Technologies Co., Ltd., Shanghai, China).
Tinción de marcación de extremos de desoxinucleotidil transferasa mediada por desoxiuridina trifosfato-biotina Nick (TUNEL)
La tinción TUNEL estaba implícita para observar la apoptosis de las células endoteliales en ratas IA sobre la base del kit TUNEL (Roche, Basilea, Suiza). Las secciones de aneurisma de rata preparadas se lavaron dos veces con xileno (5 min / tiempo) y se deshidrataron con una serie descendente de alcohol (100%, 95%, 80%, 75%) durante 3 veces (5 min / tiempo). Los tejidos se trataron con solución de proteasa K 20 μg / ml sin DNasa durante 15-30 min, se vertieron con 50 μL de solución de reacción TUNEL durante 60 min y se desarrollaron con 50 μL de diaminobencidina (DAB) a 25 ° C durante 10 min. Luego, las secciones se tiñeron por contraste con hematoxilina, se deshidrataron con alcohol en gradiente, se aclararon con xileno y se sellaron con goma neutra. Las secciones se observaron al microscopio óptico y se calculó el índice de apoptosis.
Aislamiento e identificación de células endoteliales vasculares de aneurisma
El aislamiento de células endoteliales se realizó de acuerdo con el método realizado por Boscolo et al. [dieciséis]. Los tejidos IA se seccionaron en 3 mm 2 fragmentos. Los tejidos se incubaron con colagenasa B al 0,1% / dispasa al 0,1% (Roche) durante 25 min a 37ºC. Luego, los tejidos desprendidos previamente se trituraron durante 2 min con una pipeta de 2 ml y se filtraron con un colador de 100 μm (Thermo Fisher Scientific, Rockford, IL, EE. UU.). Posteriormente, la suspensión celular se centrifugó y luego se resuspendió en medio MV2 (incluidos factores de crecimiento y suero bovino fetal al 20%) (PromoCell, Heidelberg, Alemania). Las células se sembraron a 1 × 10 4 células / mL en un matraz de cultivo recubierto con 1 μg / cm 2 fibronectina. Siguiendo el método descrito por Jackson et al. [17], las células de 80-100% de confluencia se separaron con las perlas (Dynabeads M-450 Tosylactived, Oxoid, Hampshire, Reino Unido) recubiertas con Ulex europaeus Agglutinin I (UEA) (Vector Laboratories, Ltd., Peterborough, Reino Unido) . Las células endoteliales adheridas a las perlas recubiertas de lectina se acumularon con un concentrador de partículas magnéticas y las células no conjugadas se lavaron con un medio basal. Las células positivas para UEA se resuspendieron en el medio de cultivo y se sembraron en el matraz de cultivo recubierto de fibronectina para mejorar la adhesión y la tasa de crecimiento de las células.
Las células se cultivaron en MV2 en portaobjetos de cámara recubiertos con fibronectina. Cuando la confluencia celular alcanzó el 80-100%, las células se fijaron en acetona a 4 ° C y se trataron con Triton X-100 al 1% durante 5 min y luego con albúmina de suero bovino (BSA) al 0,5% durante 15 min. Las células se gotearon con anticuerpo primario contra vWF (1:300, Abcam, Cambridge, MA, EE. UU.) Y se incubaron durante 2 h (la NC se realizó en ausencia de anticuerpo primario), se gotearon con inmunoglobulina G con conjetura de peroxidasa de rábano picante (1:150, Abcam) y se incubó durante 30 min. Luego, las células se desarrollaron con 50 μL de DAB a 25 ° C durante 5 min, se contratiñeron con hematoxilina, se diferenciaron con ácido clorhídrico al 0,1%, se deshidrataron con alcohol, seguido de aclaramiento de xileno y sellado de goma neutra. Después del secado, las células se fotografiaron bajo un microscopio invertido.
Agrupación y transfección de células
Las células endoteliales vasculares del aneurisma en la fase logarítmica se dividieron en 5 grupos:grupo blanco (células endoteliales vasculares del aneurisma sin ningún tratamiento), grupo sh-NC (células endoteliales vasculares del aneurisma transfectadas con el plásmido sh-MALAT1 NC), grupo sh-MALAT1 ( células endoteliales vasculares de aneurisma transfectadas con plásmido sh-MALAT1), grupo Oe-NC (células endoteliales vasculares de aneurisma transfectadas con plásmido NC Oe-MALAT1) y grupo Oe-MALAT1 (células endoteliales vasculares de aneurisma transfectadas con plásmido Oe-MALAT1). Los plásmidos anteriores fueron sintetizados por Genechem. La transfección celular se llevó a cabo de acuerdo con las instrucciones de la lipofectamina TM 2000 reactivo (11668-027, Invitrogen, Carlsbad, California, EE. UU.).
Ensayo de bromuro de 3- (4, 5-dimetiltiazol-2-Yl) -2, 5-difeniltetrazolio (MTT)
Las células endoteliales vasculares de cada grupo se sembraron en una placa de 96 pocillos a una densidad de 3 × 10 4 células / mL y cultivado a 37 ° C, 5% CO 2 durante 48 h. Cada grupo se configuró con 5 pocillos paralelos, y cada pocillo se añadió con 20 μL de solución de MTT fresca (5 mg / mL, Sigma, St. Louis, MO, EE. UU.). Después de 4 h de reacción, las células se mezclaron con 200 µl de dimetilsulfóxido. Después de la disolución completa, el valor de densidad óptica de las células de cada grupo se midió con un lector de microplacas (BioRad, Hercules, California, EE. UU.) A 490 nm.
Citometría de flujo
La distribución del ciclo celular se probó mediante tinción con yoduro de propidio (PI). Las células endoteliales vasculares se separaron, se centrifugaron, se resuspendieron con etanol al 75% preenfriado y se fijaron durante la noche a -20 ° C. Las células se centrifugaron para descartar el sobrenadante. Las células se agregaron en 450 μL de PBS, se agregaron con 100 μL de Rnasa A y se tiñeron con 400 μL de PI a 4 ° C durante 30 min evitando la luz. Se utilizó un citómetro de flujo (FACSCalibur, Becton, Dickinson and Company, Nueva Jersey, EE. UU.) Para probar la distribución del ciclo celular.
La apoptosis celular se ensayó mediante doble tinción con Anexina V / PI. Las células endoteliales desprendidas se recogieron y lavaron con PBS 3 veces. Las células se resuspendieron con 100 μl de tampón de unión 1 × preenfriado y se mezclaron con 5 μl de anexina y 5 μl de PI, respectivamente. La apoptosis celular se probó mediante un citómetro de flujo. Con AnnexinV como eje transversal y PI como eje longitudinal, el cuadrante superior izquierdo representaba células mecánicas lesionadas (AnnexinV - / PI + ), el cuadrante superior derecho para células apoptóticas tardías o células necróticas (AnnexinV + / PI + ), el cuadrante inferior izquierdo para células normales negativas (AnnexinV - / PI - ) y el cuadrante inferior derecho para las células apoptóticas tempranas (AnnexinV + / PI - ). Las células apoptóticas totales (AnnexinV + / PI - y AnnexinV + / PI + ) se calcularon y expresaron como un porcentaje.
Reacción en cadena cuantitativa de la polimerasa con transcripción inversa (RT-qPCR)
Trizol (Takara Biotechnology Ltd., Dalian, China) extrajo los ARN totales en los tejidos y las células y se determinó la concentración y pureza del ARN. El proceso de transcripción inversa de ARN en ADN complementario se llevó a cabo de acuerdo con las instrucciones del kit de transcripción inversa (K1621, Fermentas, Maryland, NY, EE. UU.). Las secuencias de cebadores MALAT1, miR-143 y VEGFA (Tabla 1) fueron diseñadas y compuestas por Genechem. Para evaluar la expresión de lncRNA, miRNA o mRNA, se realizó RT-qPCR utilizando SYBR GreenPCR Master Mix (Takara, Tokio, Japón) con el sistema Roche Real-Time PCR (Roche). U6 se estableció como un parámetro interno de miR-143, mientras que MALAT1 y VEGFA, con gliceraldehído-3-fosfato deshidrogenasa (GAPDH) como parámetros internos. Los niveles de transcripción relativos de los genes diana se calcularon mediante 2 - △△ Ct método.
Análisis de Western Blot
Se extrajo la proteína total de tejidos y células. La concentración de proteína se determinó mediante las instrucciones del kit de ácido bicinconínico (Boster Biological Technology Co. Ltd., Wuhan, Hubei, China). La proteína se separó por electroforesis con gel de poliacrilamida al 10% (Boster Biological Technology). La membrana se transfirió a una membrana de fluoruro de polivinilideno y luego se selló con BSA al 5% durante 1 h. La membrana se incubó con anticuerpo primario contra Ki-67 (1:1000), VEGFA (1:1000), vWF (1:1000) y metaloproteinasa de matriz (MMP) -9 (1:1000, Abcam, Cambridge, Reino Unido). , Ciclina D1 (1:1000), Bax (1:1000) y Bcl-2 (1:1000, Santa Cruz Biotechnology, Santa Cruz, California, EE. UU.) Y GAPDH (1:2000, Jackson Immuno Research, Grove, Pennsylvania, EE. UU.) Y con el anticuerpo secundario (1:500, Jackson Immuno Research) marcado con peróxido de rábano picante. La membrana se obtuvo mediante el sistema de escaneo de imágenes por fluorescencia infrarroja de dos colores Odyssey, y los valores de gris de las bandas se midieron con el software de análisis de imágenes Cantidad Uno.
Ensayo génico indicador de luciferasa dual
Los sitios de unión de MALAT1 y miR-143 fueron pronosticados y expuestos por el sitio web de bioinformática (https://cm.jefferson.edu/rna22/Precomputed/). La relación de unión entre MALAT1 y miR-143 se verificó adicionalmente mediante el ensayo del gen indicador de luciferasa. El fragmento sintético del gen de la región 3 'no traducida de MALAT1 (3'UTR) se introdujo en los vectores indicadores de luciferasa pmiR-Report (Thermo Fisher Scientific) para generar MALAT1 de tipo salvaje (MALAT1-WT) mediante el sitio de endonucleasa Bamh1 y Ecor1. El sitio de mutación de la secuencia complementaria de la secuencia se diseñó en MALAT1-WT, y el fragmento diana se insertó en los vectores informadores de luciferasa pmiR-Report para producir el tipo mutante MALAT1 (MALAT1-MUT) mediante ADN ligasa T4 después de la digestión con endonucleasas de restricción. El MALAT1-WT y MALAT1-MUT correctamente secuenciados se cotransfectaron con el imitador NC y el imitador de miR-143 en células endoteliales vasculares. Las células se recolectaron y lisaron 48 h después de la transfección, y la actividad de luciferasa se evaluó mediante un kit de detección de luciferasa (BioVision, San Francisco, CA, EE. UU.) Con un luminómetro (Glomax20 / 20, Promega, Madison, Wisconsin, EE. UU.).
La relación objetivo de miR-143 y VEGFA y el sitio de unión de miR-143 y VEGFA 3'UTR se pronosticaron a través del sitio web de bioinformática (http://www.targetscan.org/vert_72/). La secuencia de la región promotora 3'UTR de VEGFA que contiene el sitio de unión de miR-143 se combinó y se clonó en vectores indicadores de luciferasa pmiR-Report para generar VEGFA-WT. Sobre la base de este informador, el sitio de unión de VEGFA-WT y miR-143 se mutó para formar VEGFA-MUT. El indicador VEGFA-WT o VEGFA-MUT se mezcló con el imitador NC o el miR-143 y luego se cotransfectó en células endoteliales vasculares durante 48 h. Después de eso, las células se lisaron y la actividad luciferasa se probó con un kit de detección de luciferasa.
Ensayo de extracción de ARN
Para verificar la relación de unión entre miR-143 y MALAT1, se implementó un ensayo de extracción de ARN. Se diseñaron tres secuencias de miARN marcadas con biotina Bio-miR-143-WT, Bio-miR-143-MUT y Bio-miR-NC y se confiaron a GenePharma Company (Shanghai, China). Estos oligonucleótidos biotinilados se transfectaron en células durante 48 h. Luego, las células se recolectaron y se incubaron con un lisado celular específico (Ambion, Austin, Texas, EE. UU.) Durante 10 min. Después de eso, el lisado se incubó con perlas de estreptavidina M-280 prerrevestidas con ARNt de levadura y sin ARNasa (todo de Sigma) a 4 ° C durante 3 h, luego se limpió dos veces con una solución de lisis fría, tres veces con una tampón salino y una vez con una solución tampón con alto contenido de sal. Se estableció una sonda de miR-143 antagonista como NC. Se extrajo el ARN total con Trizol y luego se evaluó el nivel de enriquecimiento de MALAT1 usando RT-qPCR.
Análisis estadístico
Todos los datos fueron expuestos por el software SPSS 21.0 (IBM Corp. Armonk, NY, EE. UU.). Los datos de medición fueron representados por media ± desviación estándar. Las comparaciones entre dos grupos se realizaron mediante una muestra independiente t prueba, mientras que las comparaciones entre múltiples grupos se evaluaron mediante análisis de varianza unidireccional (ANOVA), y la comparación por pares se implementó mediante la prueba de comparaciones múltiples de Tukey. La relación entre la expresión de MALAT1 y las características clínico-patológicas de IA se determinó mediante la prueba de chi-cuadrado. P un valor inferior a 0,05 indica una diferencia estadísticamente significativa.
Resultados
MALAT1 y VEGFA están sobreexpresados y miR-143 está regulado negativamente en tejidos IA
La expresión de ET-1 y vWF en suero en el grupo IA y el grupo de control se detectaron mediante ELISA, y los resultados manifestaron que la expresión de ET-1 y vWF aumentó en el grupo IA frente al grupo de control (ambos P <0.05) (Fig. 1a, b).
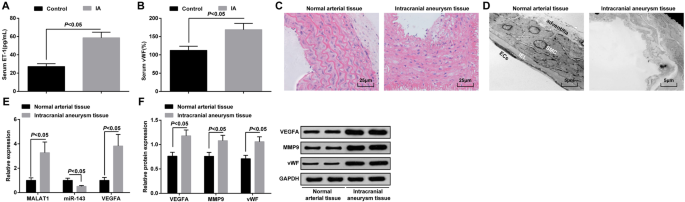
MALAT1 y VEGFA están sobreexpresados y miR-143 está regulado negativamente en los tejidos IA. un Expresión de ET-1 en suero de pacientes IA y pacientes con epilepsia del lóbulo temporal mediante ELISA. b Expresión de vWF en suero de pacientes IA y pacientes con epilepsia del lóbulo temporal mediante ELISA. c Observación patológica de tejidos IA y tejidos arteriales normales mediante tinción HE. d Observación morfológica de tejidos IA y tejidos arteriales normales mediante un microscopio electrónico de transmisión. e Expresión de ARNm de MALAT1, miR-143 y VEGFA en tejidos IA y tejidos arteriales normales mediante RT-qPCR. f Expresión de proteínas de VEGFA, MMP-9 y vWF en tejidos IA y tejidos arteriales normales mediante análisis de transferencia Western. Células endoteliales (CE), lámina elástica interna (IEL), células de músculo liso (SMC). Los datos de medición se representaron como media ± desviación estándar; las comparaciones entre grupos se realizaron mediante una muestra independiente t prueba
Los cambios patológicos de los tejidos IA se observaron mediante tinción HE. No se observaron daños evidentes en el endodermo, las células endoteliales y las células del músculo liso en los tejidos arteriales normales, y las células estaban ordenadas y tenían una estructura completa. Los tejidos IA presentaban células endoteliales dañadas, células de músculo liso degeneradas, pared arterial atenuada, fibras elásticas rotas y células inflamatorias infiltradas (fig. 1c).
Los cambios de ultraestructura de IA y tejidos arteriales normales se observaron mediante un microscopio electrónico de transmisión. Se encontró que las células endoteliales estaban completas y la estructura de la adventicia estaba intacta; no se observaron membranas elásticas internas rotas o células de músculo liso apoptóticas en los tejidos arteriales normales. En los tejidos IA, se presentó con una degeneración severa de la pared de los vasos sanguíneos, principalmente manifestada como la desaparición de la mayoría de las células endoteliales, la capa elástica interna severamente rota, las células del músculo liso severamente dañadas y degradadas, y el desorden de la membrana externa del vaso sanguíneo. (Figura 1d).
Se realizó RT-qPCR para determinar la expresión de ARNm de MALAT1, miR-143 y VEGFA, mientras que se realizó un análisis de transferencia de Western para la expresión de proteínas de VEGFA, MMP-9 y vWF en tejidos IA y tejidos arteriales normales. Se demostró que, a diferencia de los tejidos arteriales normales, los niveles de expresión de MALAT1, VEGFA, MMP-9 y vWF aumentaron y la expresión de miR-143 se degradó en los tejidos IA (todos P <0.05) (Fig. 1e, f).
El grado de Hunt-Hess, el grado de daño endotelial y el historial de tabaquismo se correlacionan con la expresión de MALAT1 en los tejidos IA
A la luz de la expresión mediana de MALAT1, los pacientes se asignaron en dos grupos:grupo de baja expresión y grupo de alta expresión. Se analizó la relación entre la expresión de MALAT1 y las características clínico-patológicas de los pacientes con IA. Los resultados sugirieron que el grado de Hunt-Hess, el grado de daño endotelial y el historial de tabaquismo se asociaron con la expresión de MALAT1 (todos P <0,05), mientras que la edad, el sexo y el modo quirúrgico no se relacionaron con la expresión de MALAT1 (todos P > 0.05) (Tabla 2).
MALAT1 regulado a la baja representa la presión arterial, la expresión de ET-1, vWF y MMP-9, así como el índice apoptótico de células endoteliales vasculares de ratas con IA
Como se muestra en la Tabla 3, la caída de MALAT1 se degradó, mientras que la restauración de MALAT1 elevó la presión arterial en la cuarta y duodécima semana.
ELISA reveló que la regulación a la baja de MALAT1 se redujo, mientras que la regulación al alza de MALAT1 elevó la expresión de ET-1 y vWF en el suero de ratas con IA (Fig. 2a, b).
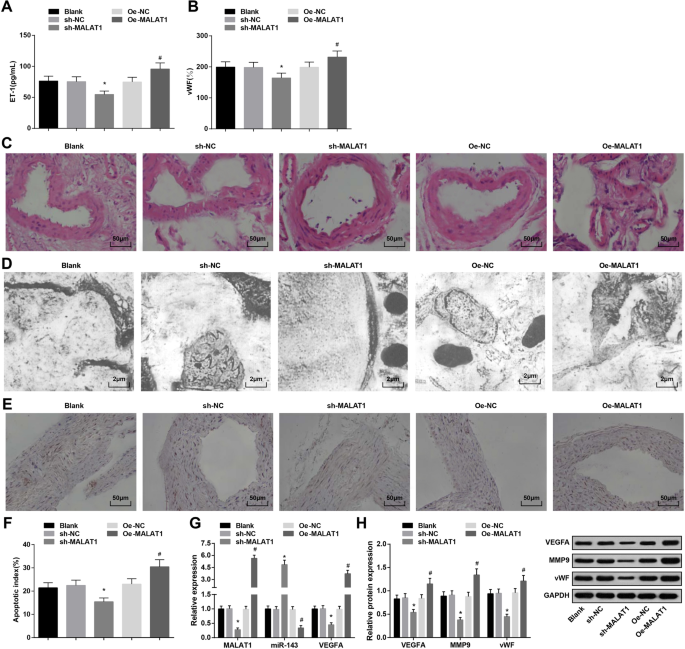
MALAT1 regulado negativamente reprime la presión arterial, la expresión de ET-1, vWF y MMP-9, así como el índice apoptótico de las células endoteliales vasculares de ratas con IA. un Expresión de ET-1 en suero de ratas mediante ELISA. b Expresión de vWF en suero de ratas mediante ELISA. c Cambios patológicos de los tejidos IA en ratas observados por tinción HE. d La ultraestructura de los tejidos IA en ratas observada por un microscopio electrónico de transmisión. e Apoptosis de células endoteliales vasculares por tinción TUNEL. f Índice apoptótico de células endoteliales vasculares de ratas. g Expresión de ARNm de MALAT1, miR-143 y VEGFA en tejidos IA de ratas mediante RT-qPCR. h Expresión de proteínas de VEGFA, MMP-9 y vWF en tejidos IA de ratas mediante análisis de transferencia Western. * P <0.05 frente al grupo sh-NC, # P <0,05 frente al grupo Oe-NC. Los datos de medición se representaron como media ± desviación estándar, y las comparaciones entre varios grupos se evaluaron mediante un análisis de varianza unidireccional seguido de la prueba de comparaciones múltiples de Tukey
Los cambios patológicos de los tejidos IA en cada grupo se estudiaron con tinción HE. El grupo en blanco, el grupo sh-NC y el grupo Oe-NC se manifestaron con una membrana interna dañada, células endoteliales exfoliadas, células musculares lisas degeneradas, células y capas reducidas y pared arterial adelgazada. En el grupo sh-MALAT1, la endodermis, las células endoteliales, la capa de células del músculo liso y la capa de la membrana externa de la arteria intracraneal estaban ligeramente dañadas pero ordenadas. El grupo Oe-MALAT1 mostró con capa íntima desaparecida, células endoteliales exfoliadas, fibras elásticas rotas y células inflamatorias infiltradas (Fig. 2c).
Los tejidos IA de las ratas de cada grupo se observaron mediante un microscopio electrónico de transmisión. Se mostró que las células endoteliales desnaturalizadas, la capa subendotelial desintegrada, la capa elástica interna desaparecida y la disminución de las células del músculo liso se presentaron en el grupo en blanco, el grupo sh-NC y el grupo Oe-NC. El grupo sh-MALAT1 se demostró con células endoteliales planas, núcleo ovalado y aumento de fibras de colágeno pero sin capa elástica. El grupo Oe-MALAT1 se caracterizó por la desaparición de las células endoteliales y la capa elástica separada de la capa muscular, que se desintegró en la luz (Fig. 2d).
El índice apoptótico de células endoteliales vasculares en ratas IA se ensayó mediante tinción TUNEL. Silenciar MALAT1 redujo el índice apoptótico de las células endoteliales vasculares, mientras que MALAT1 sobreexpresado exhibió el efecto opuesto (Fig. 2e, f).
La detección de RT-qPCR de la expresión de ARNm de MALAT1, miR-143 y VEGFA, y el análisis de transferencia Western de la expresión de proteínas de VEGFA, MMP-9 y vWF en tejidos IA mostraron que la eliminación de MALAT1 deprimía la expresión de MALAT1, VEGFA, MMP-9 y vWF y expresión de miR-143 aumentada. Por el contrario, la elevación de MALAT1 impuso influencias opuestas sobre estas expresiones génicas (Fig. 2g, h).
La baja expresión de MALAT1 promueve la viabilidad y restringe la apoptosis de las células endoteliales vasculares en IA
Se empleó tinción inmunohistoquímica para detectar la expresión del marcador endotelial específico vWF. Se manifestó que el citoplasma de las células endoteliales vasculares IA estaba cubierto de finas partículas marrones, lo que resultó positivo, mientras que el citoplasma en su grupo NC no presentaba partículas marrones. Los resultados confirmaron que las células cultivadas eran células endoteliales (Fig. 3a, b).
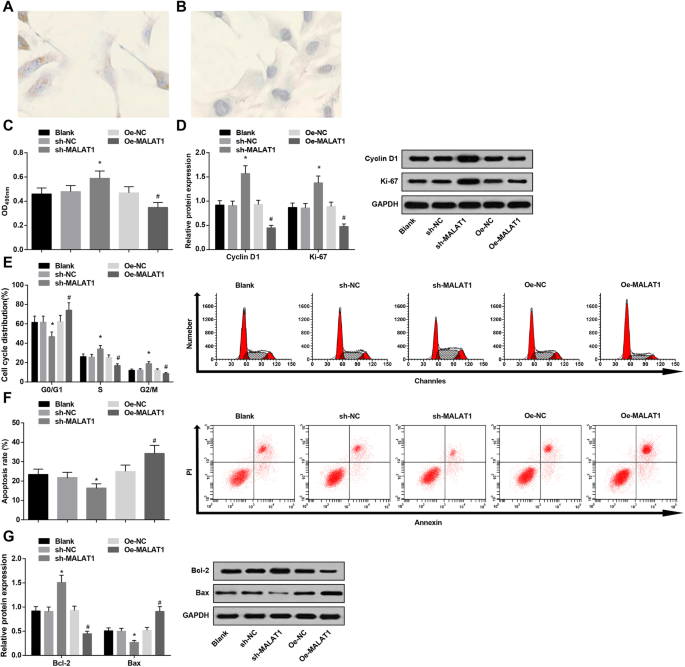
La baja expresión de MALAT1 promueve la viabilidad y restringe la apoptosis de las células endoteliales vasculares en IA. un Tinción inmunohistoquímica de vWF en células endoteliales vasculares IA:las células endoteliales vasculares IA se cubrieron con finas partículas amarillas. b Tinción inmunohistoquímica de vWF en células endoteliales vasculares IA:las células endoteliales vasculares IA no mostraron partículas marrones en el grupo NC. c Viabilidad de las células endoteliales vasculares en cada grupo mediante ensayo MTT. d Expresión de proteínas de CyclinD1 y Ki-67 en cada grupo mediante análisis de transferencia Western. e El ciclo celular cambia en cada grupo por tinción con PI. f Tasa de apoptosis celular en cada grupo mediante doble tinción con anexina V / PI. g Expresión de la proteína Bax y Bcl-2 en cada grupo mediante análisis de transferencia Western. * P <0.05 frente al grupo sh-NC, # P <0,05 frente al grupo Oe-NC. Los datos de medición se representaron como media ± desviación estándar, y las comparaciones entre varios grupos se evaluaron mediante un análisis de varianza unidireccional seguido de la prueba de comparaciones múltiples de Tukey
Se utilizaron el ensayo MTT, la citometría de flujo, junto con el análisis de transferencia Western para probar la viabilidad y la apoptosis de las células endoteliales vasculares. Se demostró que la disminución de MALAT1 promovió la viabilidad de las células endoteliales vasculares (mayor expresión de Ciclina D1 y Ki-67) y deprimió la apoptosis (disminución de células en fase G0 / G1 y aumento de células en fase S y G2 / M, reducción de Bax y expresión elevada de Bcl-2) . Sin embargo, la regulación positiva de MALAT1 funcionó de manera opuesta a la de la disminución de MALAT1 en la viabilidad celular y la apoptosis (Fig. 3c-g).
MiR-143 está unido a MALAT1 y VEGFA es un gen objetivo de miR-143
La expresión de MALA1, miR-143 y VEGFA en las células endoteliales vasculares de cada grupo se verificó mediante RT-qPCR y análisis de transferencia Western. Los resultados mostraron que la eliminación de MALA1 deprimía la expresión de MALA1 y VEGFA y mejoraba la expresión de miR-143. Por el contrario, la regulación positiva de MALA1 condujo al incremento de la expresión de MALAT1 y VEGFA y a la reducción de la expresión de miR-143 (Fig. 4a, b).
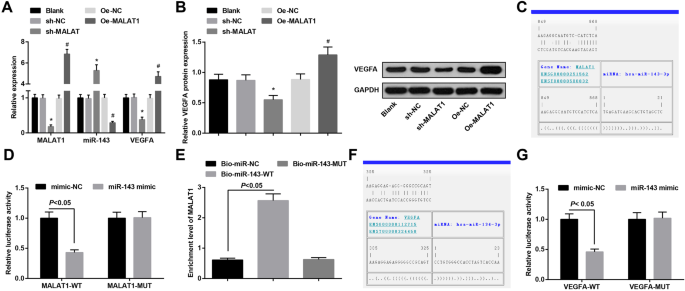
MiR-143 está unido a MALAT1 y VEGFA es un gen diana de miR-143. un Expresión de ARNm de MALAT1, miR-143 y VEGFA en células endoteliales vasculares del aneurisma en cada grupo. b Expresión de la proteína VEGFA en células endoteliales vasculares del aneurisma en cada grupo. c Los sitios de unión de MALAT1 y miR-143 predichos por el sitio web de bioinformática. d La relación reguladora de MALLA1 y miR-143 validada por ensayo de gen indicador de luciferasa dual. e La relación de unión entre MALAT1 y miR-143 verificada mediante un ensayo de extracción de ARN. f Los sitios de unión de miR-143 y VEGFA predichos por el sitio web de bioinformática. g La relación reguladora de miR-143 y VEGFA validada por ensayo de gen indicador de luciferasa dual. * P <0.05 frente al grupo sh-NC, # P <0,05 frente al grupo Oe-NC. Los datos de medición se representaron como media ± desviación estándar, las comparaciones entre dos grupos se evaluaron mediante una muestra independiente t y las comparaciones entre varios grupos se evaluaron mediante un análisis de varianza unidireccional seguido de la prueba de comparaciones múltiples de Tukey
La región de unión específica entre MALAT1 y miR-143 se adivinó mediante un software de análisis en línea (Fig. 4c). Los resultados del ensayo del gen indicador de luciferasa dual revelaron que la actividad de la luciferasa estaba alterada en el grupo imitador MALAT1-WT + miR-143 frente al grupo MALAT1-WT + imitador-NC ( P <0,05). Sin embargo, no hubo una diferencia clara en la actividad de luciferasa en el grupo imitador MALAT1-MUT + miR-143 en relación con el grupo MALAT1-MUT + imitador-NC ( P > 0,05), lo que indica que miR-143 se unió específicamente a MALAT1 (Fig. 4d). Los resultados del ensayo de extracción de ARN informaron que el nivel de enriquecimiento de MALAT1 en el grupo Bio-miR-143-WT aumentó en comparación con el grupo Bio-miR-NC ( P <0.05), pero no hubo una diferencia clara en el nivel de enriquecimiento de MALAT1 en el grupo Bio-miR-143-MUT ( P > 0.05) (Fig. 4e).
El software de bioinformática adivinó una relación específica entre miR-143 y VEGFA (Fig. 4f). Los resultados de la actividad luciferasa mostraron que la actividad luciferasa relativa reprimida después de que VEGFA-WT y miR-143 imitan cotransfectadas en células endoteliales vasculares ( P <0,05). Sin embargo, la actividad luciferasa relativa de las células endoteliales vasculares no se vio afectada por la cotransfección de VEGFA-MUT y miR-143-mimic ( P > 0,05) (figura 4g). Se indicó que VEGFA era un gen diana directo de miR-143.
Discusión
La AI es una enfermedad intracraneal grave, que conduce principalmente a hemorragia subaracnoidea [18]. Un estudio anterior ha demostrado la participación de las redes de ARN endógeno competitivas relacionadas con lncRNA en IA [19]. Además, un estudio reciente ha proporcionado una prueba de que el polimorfismo funcional en la región promotora del gen miR-143/145 está relacionado con el riesgo de IA [20]. En un estudio realizado por Xu et al . , se muestra que la sobreexpresión de factores angiogénicos, como VEGFA, puede estar relacionada con la formación y rotura de IA [21]. Para explicar el mecanismo molecular de MALAT1 en IA, se han realizado una serie de ensayos y los resultados revelaron que IA inducida por lesión endotelial vascular estaba regulada por el eje de señal MALAT1 / miR-143 / VEGFA.
En primer lugar, nuestro estudio ha proporcionado pruebas sustanciales de que MALAT1 y VEGFA están regulados positivamente y miR-143 está regulado negativamente en los tejidos IA. Un estudio reciente ha presentado que MALAT1 es uno de los lncRNAs más regulados al alza en el proceso de isquemia cerebral [22]. Otro estudio ha presentado que MALAT1 está regulado positivamente en las células de cáncer de ovario y tiene la intención de participar en los procesos de apoptosis, migración y proliferación de las células de cáncer de ovario [23]. Un estudio sobre el perfil de expresión de la AI no rota y rota ha demostrado que la expresión de factores angiogénicos como VEGFA está regulada al alza en el aneurisma roto [21]. Además, un estudio clínico ha demostrado que el grupo miR-143/145 de pacientes con AI está regulado a la baja en comparación con los sujetos sanos [11]. Además, se discutió previamente que miR-143/145 participa en varios procesos biológicos asociados con la formación de aneurismas y está regulado a la baja en pacientes con IA [20]. Todos estos hallazgos se repiten más o menos con los resultados de la exploración anterior.
Excepto por los hallazgos mencionados anteriormente, este estudio también ha explorado el papel funcional de MALAT1 en IA a través de ensayos de pérdida de función y ganancia de función. Se podría resumir que la regulación a la baja de MALAT1 redujo la presión arterial, los niveles séricos de ET-1 y la expresión de vWF y MMP-9 en tejidos IA. Se ha sugerido anteriormente que la regulación a la baja de MALAT1 restringe la regulación al alza del producto de transcripción ET-1 inducido por glucosa [24]. Además, se ha informado de que la expresión ectópica de MALAT1 es el inductor de la generación de vWF [25]. Otro estudio ha verificado que el agotamiento de MALAT1 en macrófagos derivados de la médula ósea inhibe la expresión de MMP-9 [26].
Además, se han realizado experimentos celulares para una mayor confirmación de las funciones de MALAT1 en IA. Los resultados han sugerido que la caída de MALAT1 promovió la viabilidad de las células endoteliales vasculares y deprimió la apoptosis en IA. De manera similar, se ha documentado que la alteración de MALAT1 mejora la arquitectura mural aórtica y retrasa el crecimiento del aneurisma [8]. Como complemento de los hallazgos de nuestro estudio, hay investigaciones que destacan que la mala expresión de MALAT1 induce la apoptosis y frena la proliferación de células de leucemia mieloide aguda [27]. Otro estudio también ha demostrado los efectos inhibidores de la caída de MALAT1 sobre la proliferación de las células del cartílago de la osteoartritis humana [28]. Además de eso, una investigación previa generalmente confirma que la regulación a la baja de MALAT1 puede inducir la apoptosis y atenuar la proliferación de células de glioma [29]. Además, la baja expresión de MALAT1 inducida por la interferencia de ARN promueve la apoptosis y suprime la proliferación de células de mieloma múltiple [30]. En conjunto, estos estudios han explicado hasta cierto punto el mecanismo molecular de MALAT1 en IA.
Además, este estudio ha demostrado que miR-143 está unido a MALAT1 y VEGFA es un gen diana de miR-143. De manera similar, un artículo sostiene que MALAT1 se une directamente a miR-143 y suprime su expresión [10]. Zhu y col. han descubierto que MALAT1 ejerce sus funciones interactuando con miR-143 en las células del carcinoma de cuello uterino [31]. Se confirma además que MALAT1 modula indirectamente VEGFA a través de miR-200b-3p [32]. Además, otro estudio ha sugerido que miR-143-3p media el efecto de ZFPM2 en una serie de objetivos proteicos en sangre, incluido el VEGFA [33]. Sin embargo, las interacciones entre MALAT1, miR-143 y VEGFA en IA no se han discutido y necesitan más estudios.
Conclusión
A partir de estos resultados, está claro que la caída de MALAT1 deprime la apoptosis y promueve la viabilidad de las células endoteliales vasculares en IA modulando el eje miR-143 / VEGFA. La red de coexpresión sugiere la conexión entre MALAT1 y miR-143 con la participación de VEGFA. Los hallazgos de este estudio revelan parcialmente la patogenia del inicio y la progresión de la AI, y los objetivos estudiados pueden ser un punto de entrada notablemente potencial para revelar la patología de la AI desde otra perspectiva. De manera limitada, se requieren más estudios a gran escala para ilustrar de manera integral los mecanismos del eje MALAT1 / miR-143 / VEGFA en IA.
Nanomateriales
- Acoplamiento de señales
- Tubos combinados
- Sistemas de señales de voltaje
- Sistemas de señales actuales
- Introducción a las señales de CA de frecuencia mixta
- Líneas de transmisión "largas" y "cortas"
- Señal de tráfico
- Manejo de señales C++
- Enrutador CNC de 3 ejes VS Enrutador CNC de 4 ejes
- Integridad de señal y PCB
- Comunicaciones por radio basadas en átomos